- The paper introduces a metacommunity model integrating competition-colonisation trade-off, spatial autocorrelation, and disturbance to explain species coexistence.
- Numerical analyses reveal that intermediate disturbance levels and patch connectivity modulate competitive dynamics to maximize coexistence.
- The study demonstrates nonlinear, oscillatory biodiversity responses under fragmentation, challenging traditional IDH predictions.
Disentangling Interactive Effects of Anthropogenic Disturbances on Biodiversity
Model Framework
This work presents a spatially-implicit metacommunity model integrating three canonical ecological mechanisms: competition-colonisation trade-off (CCTO), the intermediate disturbance hypothesis (IDH), and spatial heterogeneity via explicit habitat autocorrelation. The authors extend Tilman's CCTO framework by introducing a flexible set of ODEs capable of incorporating habitat loss (H), spatial autocorrelation (A), and disturbance (μ), thus facilitating the analysis of their individual and joint effects on coexistence outcomes. Dispersal is modeled continuously from strictly local to global, with a novel λ parameter (Eq. 5) governing the probability of propagule landing in habitable patches as a function of d, H, and A.
The model implements a strict competitive hierarchy, but can be extended to an arbitrary number of species, all with parameterized colonisation and dispersal rates, thus allowing gradient-based and oscillatory coexistence analyses. Analytical results are provided for two species and recursions exist for N-species competitive hierarchies.
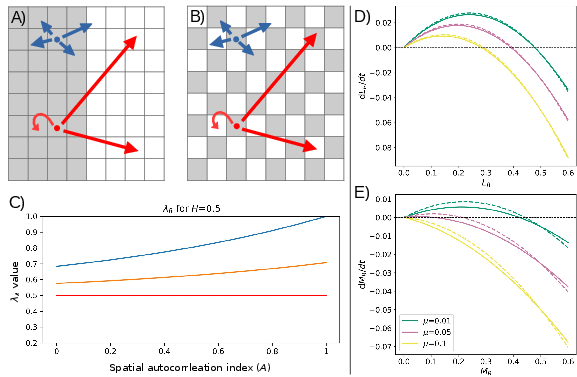
Figure 1: Schematic of model: spatial landscapes, dispersal strategies, and how autocorrelation and dispersal jointly regulate λ as a function of patch configuration and fragmentation.
Influence of Habitat Loss, Disturbance, and Spatial Autocorrelation
Numerical solutions with two and multiple species show that habitat loss, disturbance, and spatial autocorrelation have highly contingent and interactive effects on community structure. Under the CCTO, increasing habitat loss (H) generally shifts dominance towards colonisers, but this effect is modulated by disturbance intensity (A0): the broadest coexistence occurs at intermediate A1, and for high A2 only low A3 allows survival of any species. Presence of spatial autocorrelation (A4) allows competitors (with low dispersal) to persist at higher A5, but the effect of A6 is highly context dependent.
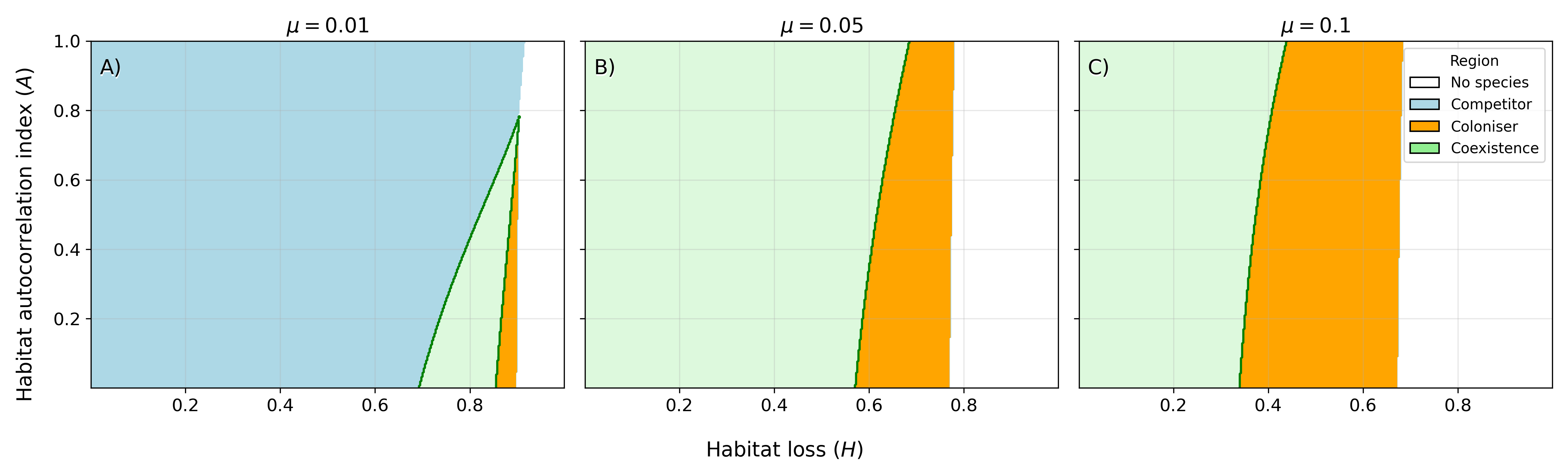
Figure 2: Coexistence regimes for coloniser and competitor across gradients of A7, A8, and A9; coexistence is maximized at intermediate disturbance and moderate spatial autocorrelation.
Spatial autocorrelation has a negligible effect without habitat loss but becomes influential when fragmented patches are present. Notably, for high μ0, competitors are advantaged, but at low μ1 and intermediate-high μ2, colonisers can dominate even with lower competitive ability.
Multi-Species Patterns: Oscillations and Non-Canonical Diversity-Disturbance Relationships
Extending the model to multiple species reveals that the interplay of disturbance, fragmentation, and spatial autocorrelation generates non-unimodal, often oscillatory, diversity-environment relationships. These include multi-peaked (bimodal or oscillatory) diversity-disturbance and diversity-fragmentation curves, in contradiction to the classical IDH paradigm.
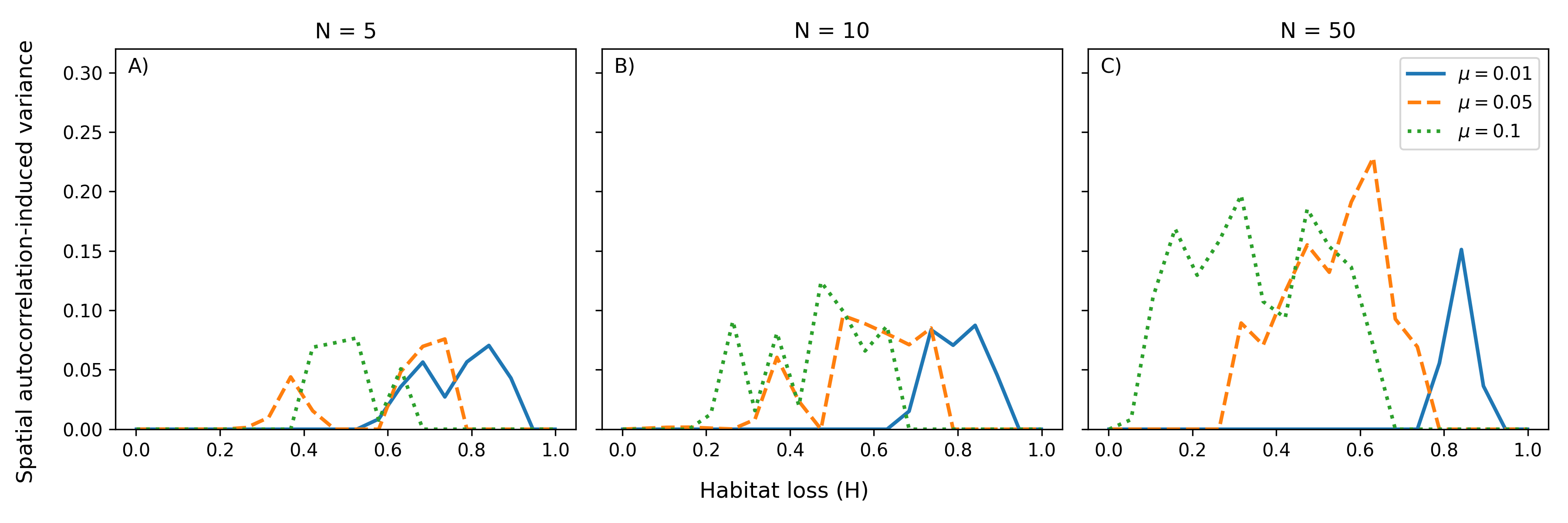
Figure 3: Spatial autocorrelation-induced variance in effective diversity as a function of habitat loss and disturbance rate for communities of increasing species richness.
Oscillatory patterns result from subsets of the competitive hierarchy transiently coexisting as environmental filters sequentially exclude or permit species along the colonisation-competition spectrum. As species number increases, the frequency of oscillations rises, and the region of parameter space supporting high diversity expands.
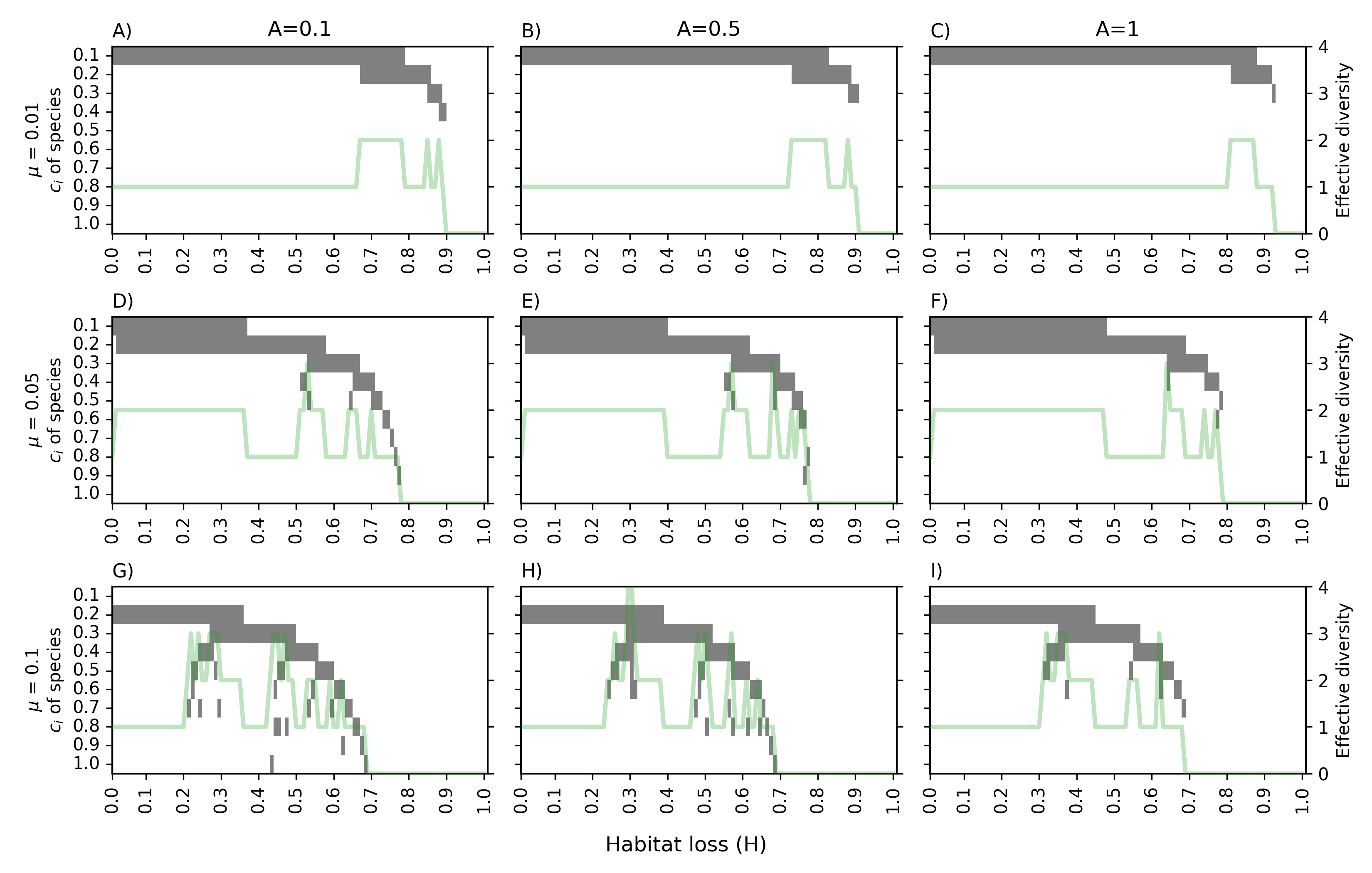
Figure 4: Co-occurrence matrices and effective diversity for 10 species; grey bands show species persistence along μ3, green line shows community effective diversity.
The diversity–disturbance relationship is highly parameter sensitive. With increasing fragmentation or decreased spatial autocorrelation, the classical unimodal IDH curve is replaced by multi-peaked or non-monotonic responses. Such shifts are accentuated with increasing species pool size and under intermediate disturbance levels.
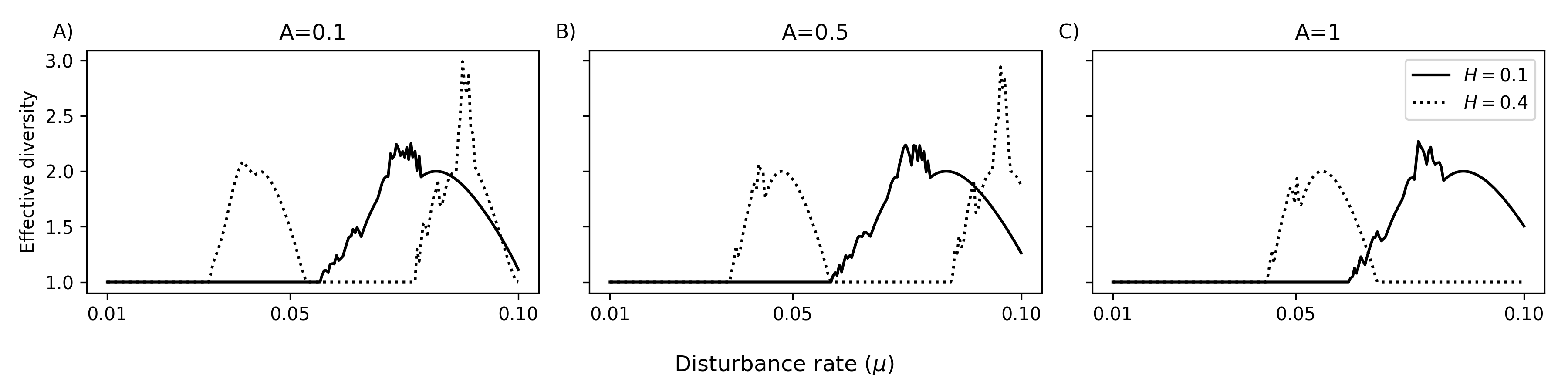
Figure 5: Diversity-disturbance relationships for 10 species; line styles indicate different μ4, panels show μ5. Patterns deviate from classical unimodal forms with fragmentation or autocorrelation.
Parameter Sensitivity and Biological Implications
The model predicts that for high diversity (large μ6) and strong fragmentation, diversity outcomes are highly sensitive to the spatial configuration of fragments and stochastic disturbance. In landscapes with intermediate-high habitat loss (μ7), spatial autocorrelation substantially augments diversity by effectively partitioning the landscape in ways that favor both local competitors and global colonisers, depending on their dispersal-dominated strategies.
For empirical studies, this model demonstrates that discrete sampling or unstratified sampling across disturbance or fragmentation gradients may yield fundamentally different biodiversity-disturbance patterns—explaining why evidence for the IDH or CCTO is inconsistent across systems. Nonlinearities and oscillations are emergent, not artifacts of fine-scale model structure.
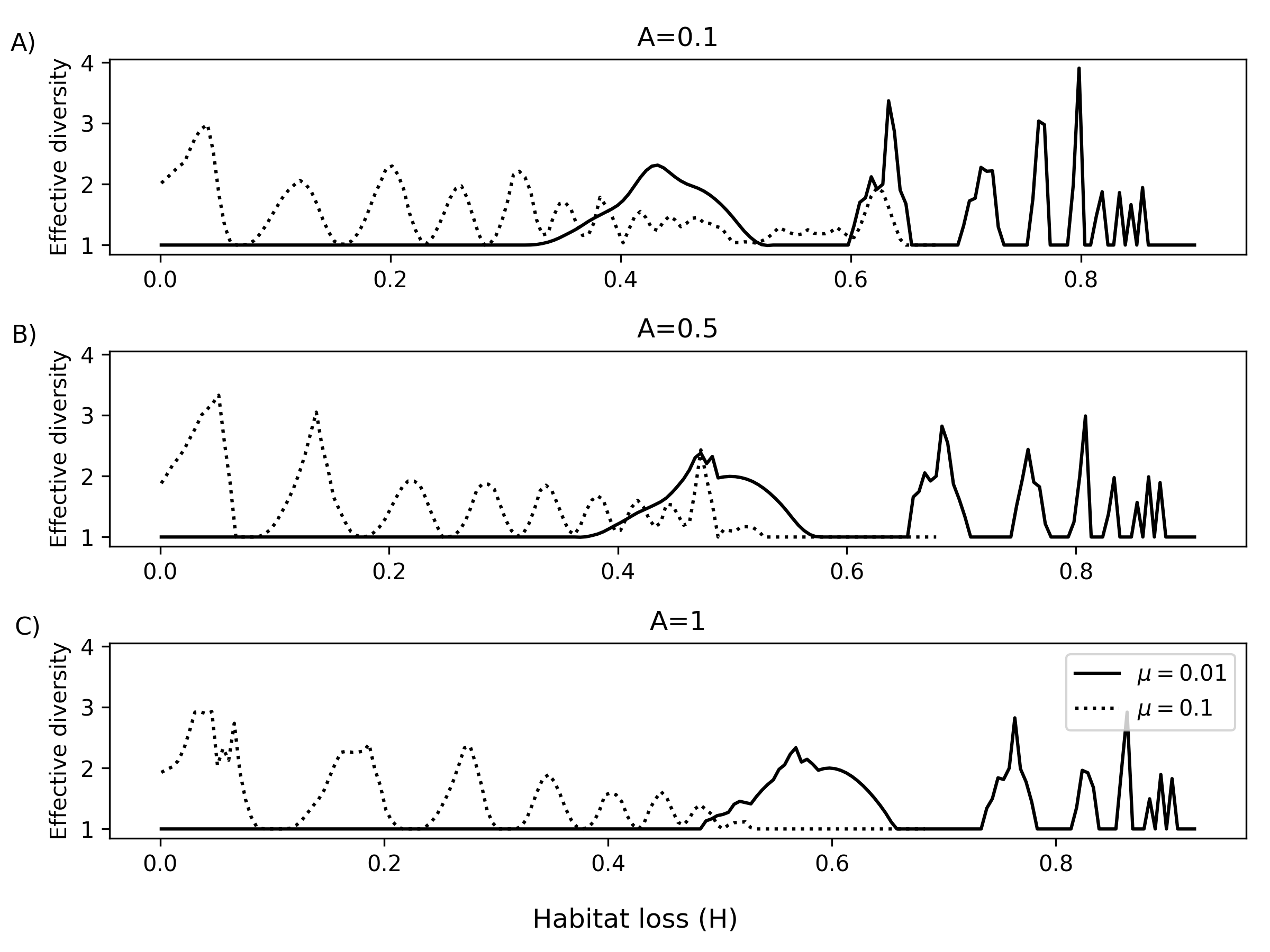
Figure 6: Effective diversity for 50 species as a function of μ8, with disturbance-level and autocorrelation cross-sections; oscillatory and non-monotonic diversity trends.
Theoretical and Practical Implications
The major theoretical implication is that the interactive effects of IDH, CCTO, and spatial structure cannot be inferred by superposition; nonadditive and emergent effects dominate, particularly in realistic biodiversity gradients. Theoretical models that neglect spatial autocorrelation or use only habitat amount as a fragmentation metric will systematically under- or over-estimate coexistence potential. Furthermore, the flexibility of the μ9 formalism enables extension to additional trait dimensions, non-hierarchical competition, and heterogeneous mortality regimes.
Practically, conservation policy emphasizing patch connectivity (autocorrelation) rather than just area per se is likely to enhance community-level resilience, especially in landscapes subject to anthropogenic disturbance. The work also suggests that efforts to detect IDH-like or CCTO-like patterns in field systems must take into account sampling scale, underlying community trait structure, and environmental autocorrelation to avoid misattribution of observed diversity patterns.
Future Directions
Future theory should extend these analyses to dynamically evolving landscapes, incorporate transient dynamics, and relax the assumption of strict competitive hierarchies. Empirical extensions could parameterize the model for real metacommunities and use landscape remote sensing to derive autocorrelation metrics, enabling quantitative linkage between model predictions and field biodiversity data.
The developed approach is structurally suitable for integration with diffusion and moment-based spatial ecological models, and could be naturally generalized to intransitive or trait-diverse communities.
Conclusion
This study delivers a tractable, extensible modeling framework demonstrating that biodiversity responses to anthropogenic disturbance and habitat fragmentation are governed by strong, context-dependent interactive effects of disturbance, spatial structure, and competitive dynamics. The emergence of non-unimodal, oscillatory diversity patterns underscores the necessity to consider both spatial configuration and multi-species interactions in explaining and predicting biodiversity patterns in disturbed landscapes. This has significant implications for both theoretical understanding and practical biodiversity management.

