- The paper introduces an environmental stress level (ESL) framework that integrates multiple environmental factors into tumor cell growth and survival modeling.
- The study employs a robust mathematical framework using ODEs, Bayesian inversion, and Sequential Monte Carlo calibration on liver cancer cell data.
- Analysis reveals that the ESL model, while matching traditional nutrient-based scaling under simple conditions, offers extensibility for multifactor environmental influences.
Environmental Stress Level Modeling of Tumor Cell Growth and Survival
Introduction
The paper "Environmental stress level to model tumor cell growth and survival" (2201.06985) advances mathematical modeling for in vitro tumor cell dynamics by formalizing the concept of an "environmental stress level" (ESL) as an auxiliary, immeasurable variable. This construct provides a principled framework to collectively quantify the influence of multiple environmental factors—such as oxygen, nutrient saturation, drug concentrations, and mechanical forces—on cell proliferation, death, and movement. The work rigorously contrasts the ESL-based approach with conventional rate scaling based on nutrient availability. Model calibration leverages Bayesian inversion and Sequential Monte Carlo (SMC), capitalizing on time-resolved population data from liver cancer cell cultures across varied nutrient saturations.
Mathematical Framework
The study formalizes three ODE systems:
Through theoretical investigations and mathematical analysis, all models guarantee positivity and boundedness of solutions (steady states and stability are characterized for all models), supporting biological feasibility.
Data Acquisition and Noise Modeling
Experimental calibration used fluorescence-based cell viability assays, correlating signal intensity to viable cell density, with datasets spanning five FBS concentrations. Calibration integrates multiplicative noise, validated empirically as gamma-distributed, ensuring realism in measurement uncertainty and protecting against spurious negative values.
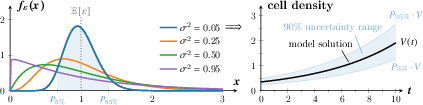
Figure 2: Left: Probability density function fε of ε∼Γ(1/σ2,1/σ2) for different σ2 and illustrative percentiles; Right: 90% uncertainty range around a sample model solution.
Bayesian Inversion and SMC Calibration
Bayesian parameter estimation is framed under comprehensive priors for all biologically relevant parameters, aligning hyperparameter choices with prior biological knowledge. SMC is employed to sample the complex posterior induced by sequential, time-resolved cell viability measurements. The process cycles through time slices and initial seeding densities, compositing evidence across 480 measurements for robust inference.
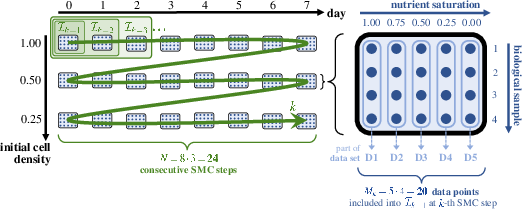
Figure 3: Schematic description of data utilization (D1–D5) for incremental SMC model calibration.
Statistical Validation and Model Selection
Calibration outputs include marginal posteriors and associated variances for all parameters. ESL model Mη introduces an additional sensitivity parameter αη, exhibiting substantial variance due to limited identifiability—primarily impacting model response in the earliest timepoints or significant environmental transitions. Quality of fit is measured using cumulative distribution-based validation metrics, complemented by evidence-based model selection (Bayes factor). The Bayes factor (log-scale) generally trends in favor of the ESL model, especially in early calibration phases, despite increased model complexity.
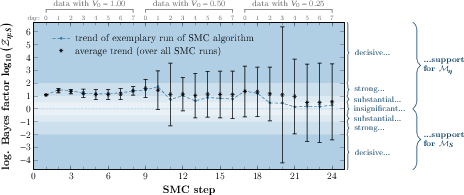
Figure 4: Logarithm of the Bayes factor Zη:S over sequential SMC calibration steps. Error bars indicate 95% confidence intervals across SMC replicates.
Numerical Results and Analysis
Both ESL-based and direct-scaling models fit the experimental data closely, with overlapping or indistinguishable parameter posteriors outside ESL-specific parameters. Deviations in calibration are primarily attributable to measurement anomalies at particular time points (t≈3 days), generating outliers and large uncertainty contributions. Model solutions remain consistent, with validation metrics approximating unity across datasets.
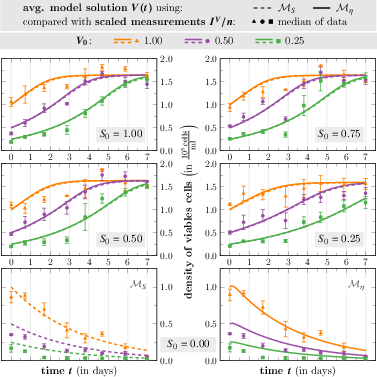
Figure 5: Time evolution of the average model solution V for each initial density V0 compared to the scaled data median.
The stress sensitivity parameter αη is found to be large, resulting in rapid attainment of steady-state η (typically within one day), effectively reducing the dynamic ESL model to steady-state scaling analogous to direct rate modulation.
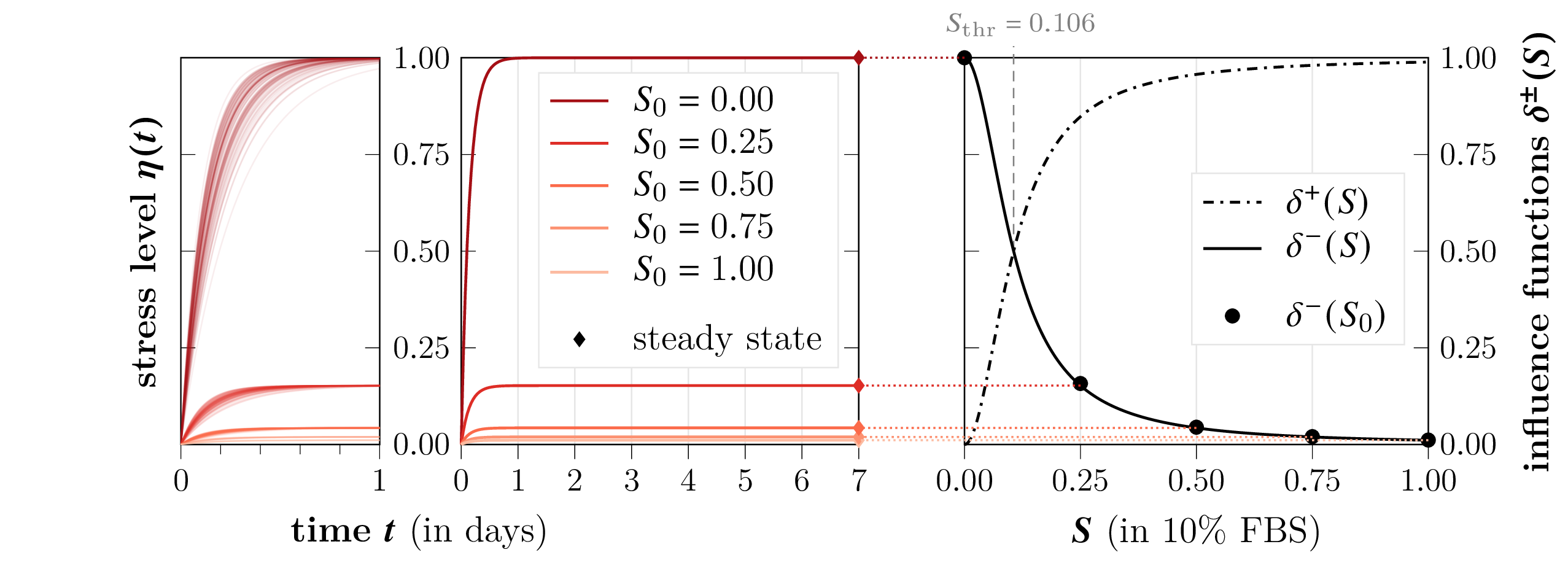
Figure 6: Time evolution of the environmental stress level η(t) for various nutrient saturations, illustrating rapid convergence to steady-state.
Correlations among model parameters are elucidated via scatter plots: strong positive correlation between proliferation and natural death rates, and moderate negative correlation between carrying capacity and logistic inhibition parameter, confirming model structural expectations.
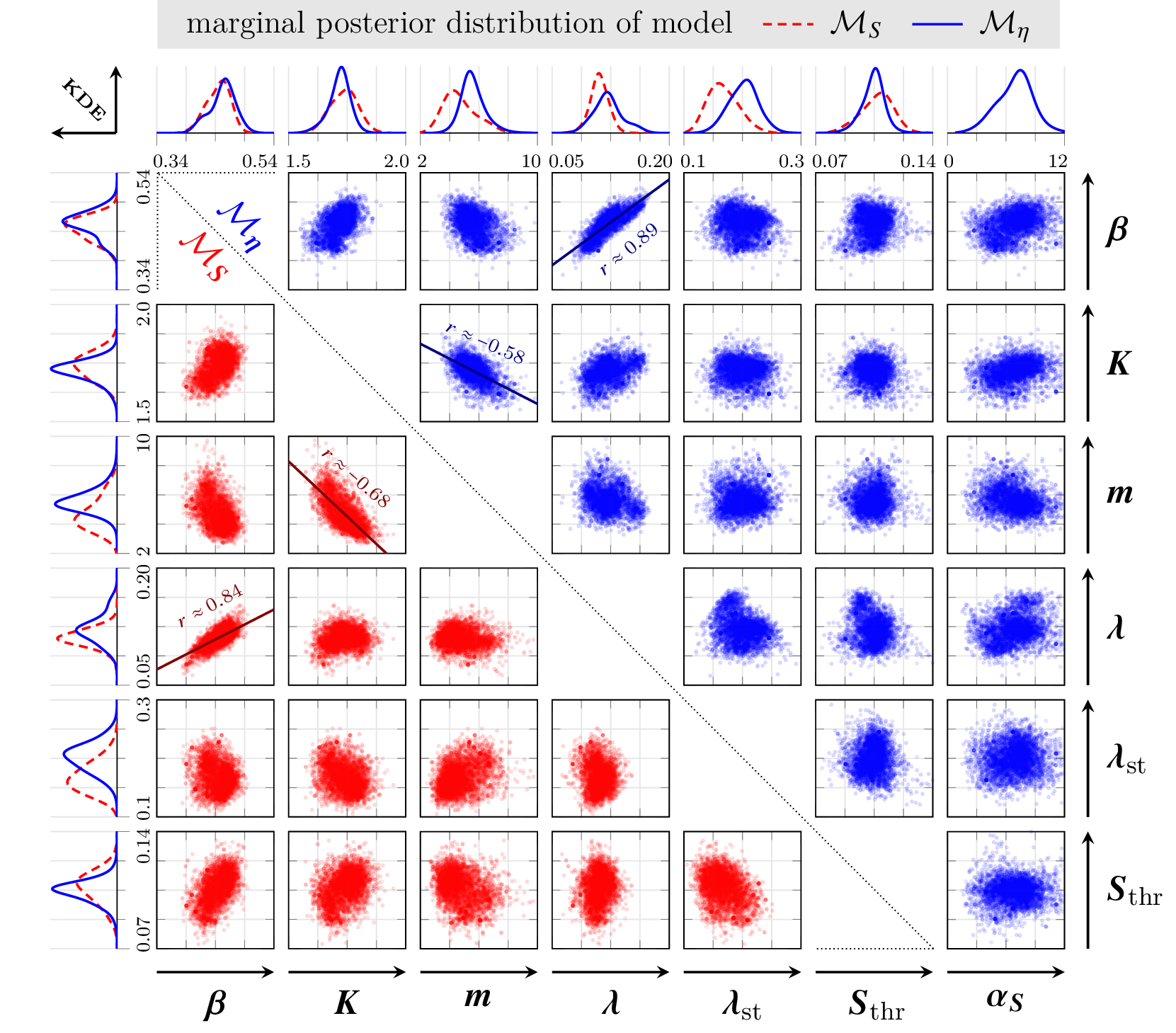
Figure 7: Marginal and pairwise posterior distributions for parameters, color-coded by model; selected pairwise scatterplots and regression lines for parameters with significant correlation.
Model Validation and Limit Analysis
Under optimal nutrient conditions, steady-state analysis and comparison with extended-duration data validate the model's reliability across growth regimes. Both ESL and direct-scaling approaches maintain predictive accuracy, though neither captures population overshoot phenomena seen in the biological data, exposing limitations inherent to logistic-based models without explicit time-lag or resource-sensing mechanisms.
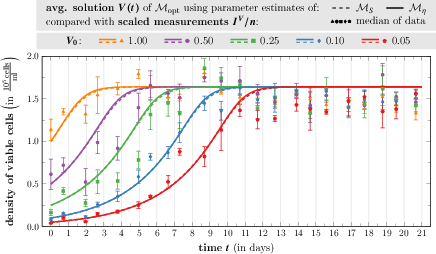
Figure 8: Time evolution of the average model solution V for the "limit model" Mopt versus median scaled validation data over 21 days.
Implications and Future Directions
The ESL framework, though only modestly superior in the current nutrient-only scenario, provides a structurally modular and extensible approach for integrating additional environmental influences—e.g., oxygen tension, drug exposure, mechanical stress—within a single variable. This enhances model interpretability and parameter inference, especially as multi-factor and spatial complexities are introduced (partial differential equation extension for motility and chemotaxis-like behaviors). The methodological infrastructure is readily adaptable to next-generation experimental systems, such as organ-on-a-chip platforms, affording realistic, controllable microenvironments for model validation and calibration.
Conclusion
The environmental stress level approach offers a mathematically rigorous, data-calibrated formalism for collective modeling of environmental influences on tumor cell dynamics. While simple experimental conditions yield nearly equivalent descriptive quality to direct scaling, the ESL stands out for theoretical and practical extensibility. As experimental sophistication grows and environmental complexity increases, the ESL methodology is poised to provide critical insights and flexible model architectures for predictive oncology and systems biology.