- The paper demonstrates that Simpson's paradox explains observed nonlinear and threshold contagion effects by aggregating heterogeneous group dynamics.
- It employs Approximate Master Equations and joint distributions of local transmission parameters to derive effective contagion kernels.
- The findings caution that neglecting group-level heterogeneity can lead to misinterpreting simple linear mechanisms as complex contagion phenomena.
Simpson's Paradox and the Emergence of Apparent Nonlinear, Threshold, and Complex Contagion Dynamics
Introduction
This work rigorously investigates the origins of observed nonlinear and threshold-like contagion processes in population-level incidence data, showing that such dynamics—commonly interpreted as genuine "complex contagion"—can frequently result from coarse-graining over underlying group-level heterogeneity and covariate correlations, rather than from intrinsic nonlinear transmission mechanisms. The authors formalize and extend the argument that Simpson's paradox fundamentally explains the ubiquity of superlinear and threshold contagion kernels when averaging across diverse subpopulations. They introduce the concept of the "Simpson's contagion," where population-level data display superlinear or complex contagion, even though every homogeneous subgroup of the population follows strictly linear or sublinear dynamics.
Theoretical Framework and Model
The population is modeled as a composition of heterogeneous groups, each characterized by two local contagion parameters: a baseline transmission rate λ and a scaling exponent ν, specifying the form of the contagion kernel β(i∣λ,ν)=λiν for group size n and i infectious members. Heterogeneity across groups is implemented via arbitrary joint distributions of (λ,ν), including structured correlations, and group dynamics are tracked using Approximate Master Equations (AMEs) that couple within-group processes to a population-level mean-field force ρ(t).
Crucially, empirical observations usually infer effective transmission kernels averaged across these groups, marginalizing out hidden group-level covariates. The authors derive closed-form and equilibrium expressions for effective kernels β~(n,i) by integrating over the heterogeneous parameter space and provide two rigorous measures of kernel variation: the population-averaged increment δβ~ and the increment within fixed-parameter groups Δβ~. Discrepancies between these offer diagnostic tools for detecting Simpson's contagions.
Case Studies: Interpreting Apparent Nonlinearity
Heterogeneous Transmission Rate (ν0) with Linear Kernel (ν1)
Populations are partitioned into groups with power-law distributed ν2 and fixed linear scaling. While each group is governed by a linear kernel, the effective kernel aggregated over the entire population exhibits a marked threshold-like nonlinearity—transmission rates shift abruptly around a critical value of ν3. Analysis shows that groups with low ν4 dominate at low prevalence, whereas those with high ν5 are overrepresented at high ν6, biasing the observed kernel.
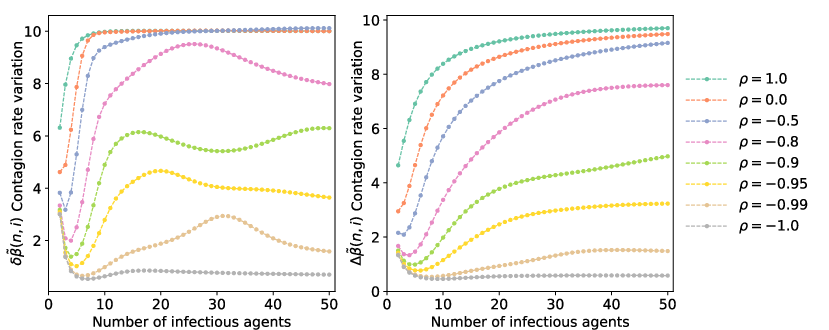
Figure 1: Percentage of Simpson's contagion as a function of correlation (parameter ν7) between ν8 and ν9, controlling the emergence of threshold and nonlinear effects.
Heterogeneous Sublinear Exponent (β(i∣λ,ν)=λiν0) with Fixed β(i∣λ,ν)=λiν1
Here, β(i∣λ,ν)=λiν2 is exponentially distributed over [0,1], making all group kernels strictly sublinear. Yet, the effective kernel, when coarse-grained, still displays a globally superlinear, threshold-like transition. For large heterogeneity (large β(i∣λ,ν)=λiν3), the system exhibits rapidly increasing returns at mid-range β(i∣λ,ν)=λiν4, then settles into population-level linearity at large β(i∣λ,ν)=λiν5. This underscores that global observations of threshold contagions can mask uniformly sublinear local dynamics.
By coupling β(i∣λ,ν)=λiν8 and β(i∣λ,ν)=λiν9 with varying correlation structures (via copula models), the authors demonstrate fine control over the shape of the effective kernel. Positive correlations (high n0 combined with high n1) amplify threshold and superlinearity, while strong negative correlations can produce alternating regimes of super- and sub-linearity or yield effective rates below the maximum of the marginal distributions.
Numerical and Analytical Results
Across all scenarios, both analytical and simulation results confirm that aggregation over unmeasured or heterogeneous group processes robustly induces nonlinear, threshold, or superlinear effective contagion kernels from purely linear or even sublinear group-level mechanisms. The magnitude and qualitative type of distortions are modulated by the heterogeneity and the correlation structure of the group-level parameters.
Implications for Modeling and Inference
These findings have significant ramifications for epidemic modeling, behavioral adoption, innovation diffusion, and inferences drawn from incidence data in complex networks. Naïve model selection or parameter inference that does not account for hidden heterogeneity or group structure will, in general, incorrectly ascribe nonlinear or threshold phenomena where none exist at the mechanistic level. The authors highlight that true identification of complex contagion should leverage:
- Local, experimental measurement of group-specific kernels;
- Statistical control for hidden covariates during parameter inference;
- Comparative analysis between population-mean kernel increments (n2) and within-group increments (n3) to diagnose Simpson contagions;
- Use of mixture models and heterogeneous effective kernels in statistical inference frameworks, instead of single kernel models, as advocated in studies of network reconstruction and community detection.
Conclusions and Future Directions
This study establishes that Simpson's paradox provides a robust, general explanation for the widespread empirical observation of nonlinear, threshold, and complex contagion processes in heterogeneous populations. The results caution against attributing apparent superlinearity or threshold phenomena to intrinsic nonlinear mechanisms without careful consideration of hidden group-level heterogeneity. Theoretically, this work motivates novel inference strategies tailored to disentangle actual complex contagion from Simpson's contagions and suggests that observed macroscopic complexity is often a statistical artifact of averaging over locally simple parts.
The analysis calls for more sophisticated data collection and model selection approaches in epidemic forecasting, social dynamics, and innovation diffusion, as well as further investigation into network structure and its interaction with contagion heterogeneity.
Figure 1: Numerical estimates of effective kernel variation n4 showcase how correlations between group-level parameters n5 and n6 drive nonlinear, threshold, or alternating patterns in observed contagion kinetics.
Conclusion
By formulating and analyzing the "Simpson's contagion," this work provides a rigorous foundation for interpreting apparent complex contagion dynamics as artifacts of marginalization over group heterogeneity and hidden covariates. Recognizing this statistical phenomenon is essential for correct inference and intervention design in complex adaptive systems. Future methodological advances should focus on robustly distinguishing true nonlinear transmission from effective nonlinearities induced by population structure.