SwitchCraft: A Programmatic Framework for Designing State-Switching Proteins
Abstract: Multistate mechanisms underlie many of the complex functions observed in natural proteins. The ability to rationally design multistate proteins would have transformative implications for many areas of biotechnology, yet lies beyond the capabilities of existing deep learning frameworks for protein design. To address this gap, we introduce SwitchCraft, a versatile and programmatic framework for designing state-switching proteins based on backpropagation through compositional design constraints parameterized by structure prediction models. In silico evaluations demonstrate success on a wide range of state-switching functional primitives, from allosteric regulation of motifs to discrimination of bound ligand identities. Using these primitives, we demonstrate an in silico strategy for de novo design of fluorescent biosensors to arbitrary small molecule analytes. These results position SwitchCraft at the inception of a powerful paradigm for higher-order functional protein design. Code is available at https://github.com/bjing2016/switchcraft.
Paper Prompts
Sign up for free to create and run prompts on this paper using GPT-5.
Top Community Prompts
Explain it Like I'm 14
What this paper is about (in simple terms)
The paper introduces SwitchCraft, a computer framework that helps scientists design proteins that can switch between different “states,” like a gadget that changes shape or behavior when a specific small molecule is present. Most current AI tools can design proteins for a single, fixed shape. SwitchCraft goes further: it lets you “program” a protein to do one thing in one situation and something different in another.
Here, a “state” means the protein’s shape and activity when certain molecules (like metals, small chemicals, or DNA) are around—or not around. Think of it like a foldable tool that rearranges itself depending on which attachment you snap on.
What questions the researchers asked
- Can we build a general, flexible way to design proteins that behave differently in different situations (multiple states)?
- Can we “turn on” or “turn off” certain protein features using small molecules, metals, or DNA (like flipping a light switch)?
- Can we design proteins that:
- switch between two different functions,
- bind to a partner only when a helper molecule is present,
- change shape depending on which molecule is bound,
- and tell similar molecules apart by adopting different shapes?
- Can we use this to build new fluorescent biosensors—proteins that light up when a specific small molecule is present?
How the method works (everyday explanation)
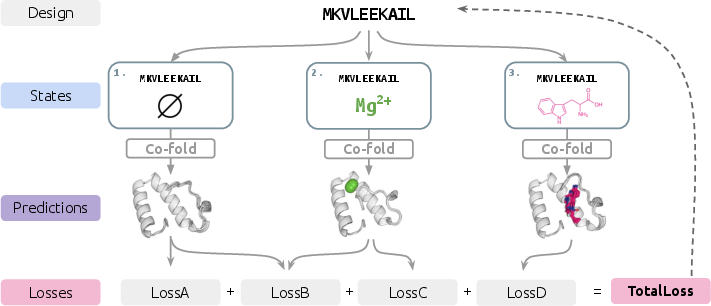
- “Programming” a protein: The team writes a set of simple rules (they call them “losses,” but think “goals”) describing what the protein should do in each state. Examples:
- “In state A, this part should hold a certain shape.”
- “In state B, this protein should bind this small molecule.”
- “The shapes in state A and state B should be clearly different.”
- Predicting shapes with AI: For a given protein sequence (the chain of amino acids—the “letters” that make the protein), a powerful structure-prediction program estimates the 3D shape it would take in each state.
- Tuning the sequence: The computer then nudges the letters of the sequence to better satisfy all the goals at once. This “nudging” uses a standard technique (backpropagation/gradient descent) that’s common in machine learning—like steadily adjusting a recipe after tasting it, so it pleases multiple judges at the same time.
- Repeat: The system loops through “predict shape → check goals → adjust sequence” many times until it finds a sequence that fits all the state-by-state rules well.
In short, SwitchCraft lets you write a “to-do list” for a protein across different conditions and then uses an AI folding model to help find a sequence that meets that list.
What they found and why it matters
The team tested SwitchCraft on several kinds of “state-switching” tasks in computer simulations (“in silico”). Below are examples they achieved:
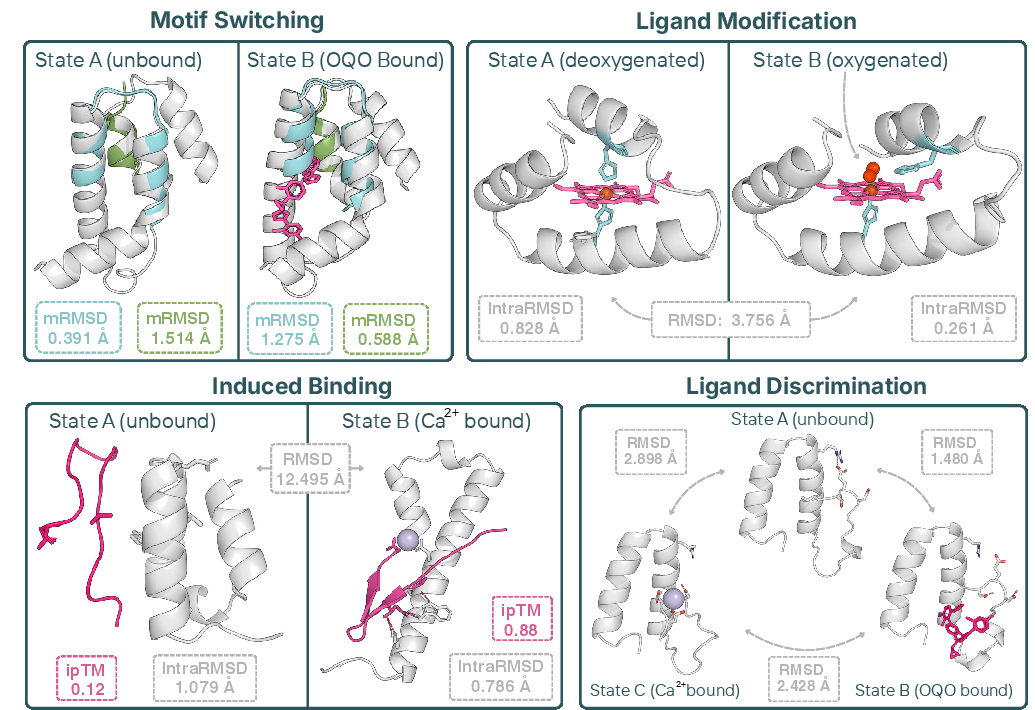
- Turning features on or off with different ligands (helpers)
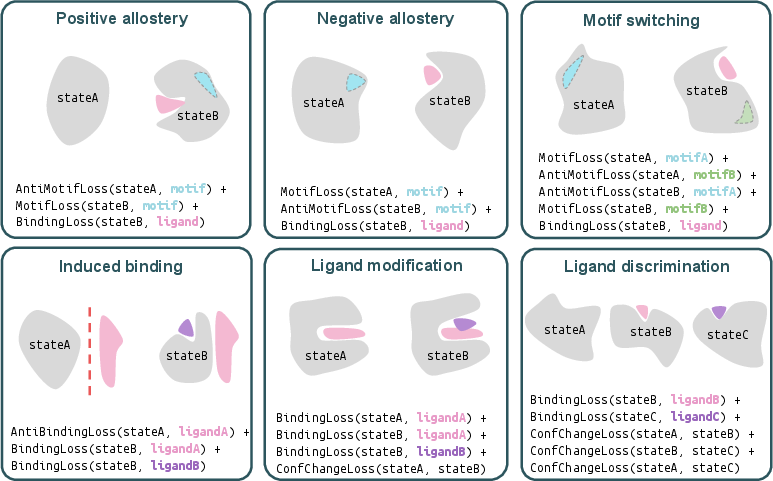
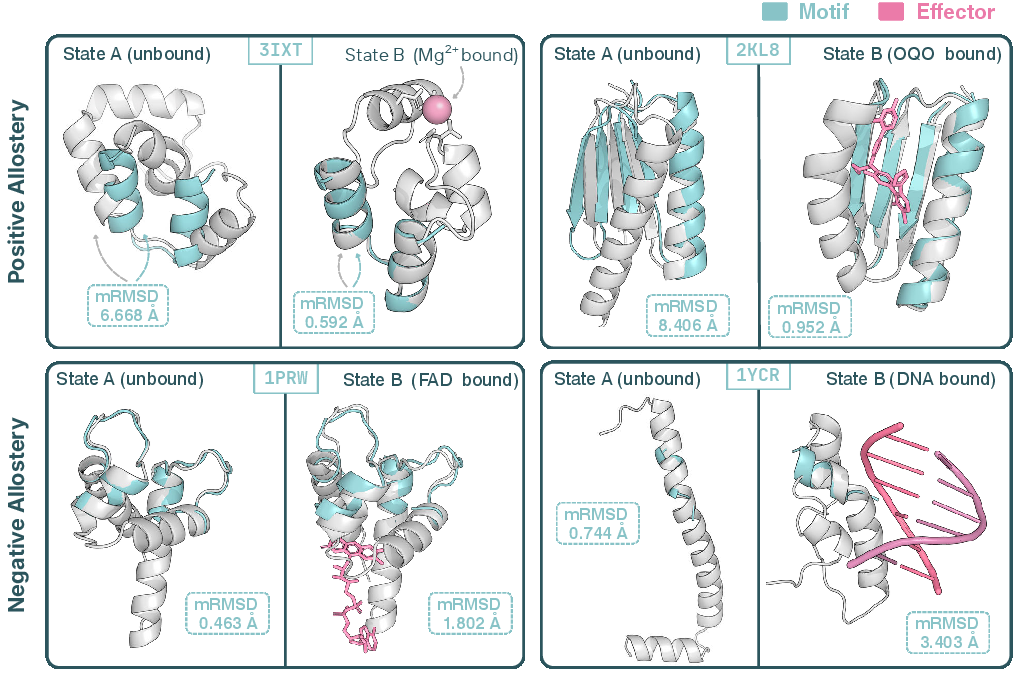
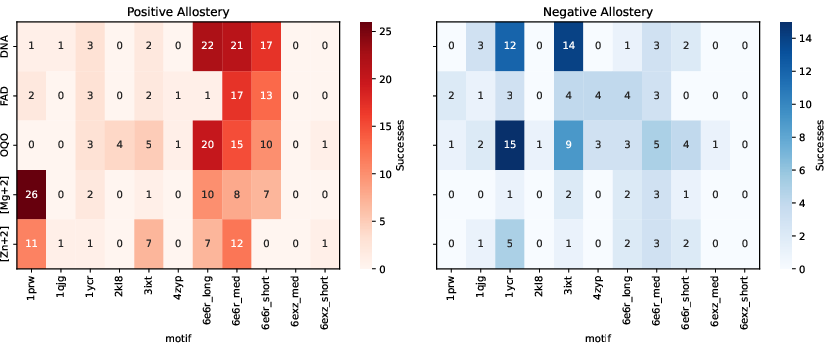
- They used small molecules, metal ions (like calcium or magnesium), and even DNA, to make a protein’s feature (called a “motif,” a small structural pattern) appear in one state and disappear in another.
- They succeeded on multiple motifs and ligands, showing both “positive allostery” (motif turns on when ligand binds) and “negative allostery” (motif turns off when ligand binds).
- Switching between two different features
- They designed proteins that show one motif without the ligand, and a different motif when the ligand is bound—like a tool that flips from one function to another depending on the attachment.
- Induced binding
- They built a protein that only grabs onto a partner peptide when calcium is present. When calcium binds, the designed protein reshapes to create the right interface for binding.
- Changing shape when a bound molecule changes
- They made a heme-binding protein predicted to shift its shape when oxygen is added—similar in spirit to how hemoglobin changes in our blood when it grabs oxygen.
- Discriminating between different ligands
- They designed a mini protein that adopts three distinct shapes: one with no ligand, one with ligand A, and another with ligand B. This shows the ability to create multi-way switches.
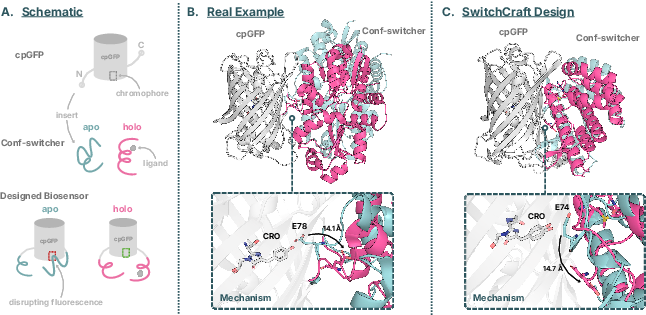
- A path to new fluorescent biosensors
- They used SwitchCraft to design “conformation switchers” for small molecules like SAM, cGMP, and ATP. Then they inserted a fluorescent protein part (cpGFP) into these switches and looked for designs predicted to brighten only when the target molecule binds.
- One design mimicked a known nicotine biosensor’s trick: in the off state, a nearby acidic amino acid dampens (quenches) the glow; when the target binds, that residue moves away, and the sensor lights up.
Why this matters: These demos show, for the first time, a general computer-based way to design proteins that can change what they do depending on their environment—something very common in nature but hard to engineer.
Note: Most results are computer predictions. The approach looks promising, but lab experiments are needed to confirm real-world function.
What this could lead to
- Smarter biosensors: Proteins that light up only in the presence of specific molecules, useful for diagnostics or tracking cell signals in real time.
- Controllable therapeutics: Proteins and binders that turn on only in certain conditions (like specific ion levels), potentially improving safety.
- Synthetic biology tools: Designer proteins that run multi-step processes, react to different inputs, or perform complex tasks—like molecular machines.
- A general “language” for protein function: As constraints get more creative, we could design ever more sophisticated, nature-inspired mechanisms (motors, switches, even multi-step enzymes).
Overall, SwitchCraft is a step toward programmable, multi-mode proteins—moving beyond single-shape designs to dynamic, responsive biomolecules that do different jobs on command.
Knowledge Gaps
Knowledge gaps, limitations, and open questions
Below is a concise list of what remains missing, uncertain, or unexplored, distilled to guide actionable follow-up research.
- Heavy reliance on a single structure-prediction model (Boltz-1) without cross-model validation
- No comparisons to AlphaFold2/OpenFold/ESMFold or ensemble agreement to assess predictor-induced bias in multistate contexts.
- Open question: how robust are designs when re-scored or re-folded by alternative predictors or physics-based methods?
- Lack of experimental validation across presented primitives and biosensor pipeline
- Aside from a brief note of preliminary work, no synthesis, expression, stability, binding, or functional switching assays are reported.
- Actionable next step: prioritize a subset of designs (e.g., induced binding, ligand discrimination, and a cpGFP-based sensor) for biochemical/biophysical validation with quantitative switching curves and dynamic range.
- Success rates are low and sample efficiency is unclear
- Many thousands of optimization trajectories yield few “successful” designs; no analysis of compute cost per success or strategies to improve yield.
- Open question: can learned proposal distributions, curriculum schedules, or multi-start/multi-armed bandit approaches reduce compute and improve success rates?
- Loss functions operate on coarse geometric proxies and may miss critical chemistry
- Motif loss is Cα/Cβ distance-based; side-chain geometry, backbone torsional preferences, and catalytic/coordination chemistries are not explicitly enforced.
- Binding loss uses thresholded distance entropy and ignores orientation, hydrogen bonding geometry, electrostatics, solvation, and desolvation energetics.
- Actionable direction: add differentiable atomic/side-chain motif constraints, directional H-bond and coordination geometry terms, and coarse electrostatics/solvation-aware terms.
- No explicit thermodynamic grounding or free-energy differences across states
- “Switching” is inferred from predictor confidence and RMSDs rather than ΔG differences between states with/without ligand.
- Open question: can approximate free-energy estimators (e.g., ML-parameterized scoring, Rosetta, GB/SA) or MD-enhanced re-scoring quantify ΔG_state and ΔΔG_switch?
- Kinetics and transition-path plausibility are not addressed
- The paper itself notes some conformational changes appear implausible without unbinding/rebinding; no path-based constraints are used.
- Actionable idea: incorporate transition-path penalties or nudged elastic band (NEB)-style differentiable constraints between states; penalize large side-chain identity changes at key interfaces that would require bond breaking/forming.
- Multi-objective loss balancing and gradient conflict are not systematically treated
- Weights (e.g., anti-binding set to −0.5) are heuristic; no adaptive weighting, gradient surgery, or Pareto-front exploration is reported.
- Open question: do automated weight schedules or multi-objective solvers improve stability and success on complex specifications?
- Generalization beyond small proteins and simple topologies is untested
- Most designs are ~50–200 AA; no results for larger, multi-domain, or flexible architectures where switching often emerges naturally.
- Actionable experiment: attempt domain-scale designs (≥300 AA) and multi-domain linkers; assess how cost and success scale with length and number of states.
- Limited ligand modeling fidelity, especially for metals, heme, DNA, and flexible small molecules
- Protonation/tautomer states, coordination numbers, oxidation states (e.g., heme), and sequence-dependent DNA mechanics are not systematically addressed.
- Actionable step: integrate cheminformatics pipelines to enumerate and co-optimize ligand protonation/tautomer states; add explicit metal coordination geometry constraints and sequence-specific DNA elastic models.
- No explicit modeling of solvent, pH, ionic strength, or membranes
- Many allosteric and binding phenomena are environment-dependent, yet the framework assumes a vacuum-like environment.
- Open question: can implicit-solvent terms or membrane-aware constraints (for GPCR-like targets) be incorporated without prohibitive cost?
- Evaluation metrics may not correlate with function
- Reliance on mRMSD thresholds, iPTM, pLDDT, intraRMSD, and crossRMSD may not predict switching thermodynamics, kinetics, or functional readouts (e.g., catalytic rate, fluorescence brightness).
- Actionable next step: benchmark correlations between these metrics and experimental outcomes; develop function-specific surrogate metrics (e.g., chromophore environment scores for fluorescence).
- No systematic ablation of losses, schedules, or STE-related biases
- Sensitivity to temperature annealing, β/γ mixing, Gumbel initialization, and STE bias is unknown.
- Actionable study: controlled ablations and hyperparameter sweeps to identify robust regimes and reduce collapse or mode-hopping across states.
- Potential overfitting to the structure predictor’s inductive biases
- Designs may exploit predictor artifacts (e.g., overconfident contacts) rather than true biophysical plausibility.
- Open question: can adversarial checks (e.g., decoy ligands, shuffled motifs) and predictor ensembles detect and reduce “model hacking”?
- Limited negative design against off-target ligands, interfaces, or oligomeric states
- Anti-binding is applied for specified contexts, but broader off-target interactions (other ligands, homodimerization/aggregation) are not discouraged.
- Actionable extension: add library-level off-target contexts during optimization (multi-ligand anti-binding, anti-oligomerization penalties, aggregation predictors).
- Biosensor pipeline lacks photophysics-aware objectives and linker optimization
- Chromophore “unquenching” is proxied by contact changes; no modeling of excited-state dynamics, pKa shifts, or brightness/spectral changes; linkers are not optimized jointly with cpGFP insertion.
- Actionable plan: integrate empirical chromophore-environment predictors, QM/MM-inspired surrogates, and differentiable linker optimization within the multistate objective.
- No standardized, open benchmark for multistate design with agreed-upon metrics
- The proposed positive/negative allostery set is a start but lacks community baselines, shared ligand parameterizations, and standardized success criteria.
- Actionable item: release curated benchmark suites (problems, ligands, evaluation scripts) and organize cross-lab evaluations.
- Scaling to >3 states and complex cycles is not demonstrated
- Only up to three states (two ligands + apo) are shown; many natural mechanisms involve long cycles and intermediate states.
- Open question: how do optimization stability and cost scale with N_states, and what strategies (e.g., curriculum over states, state sampling) preserve convergence?
- No integration with sequence-level fitness priors
- Solubility, expression, disorder/aggregation, and immunogenicity are not considered during optimization.
- Actionable extension: add PLM-based priors, solubility/aggregation predictors, and disulfide/glycosylation handling to penalize problematic sequences.
- Limited comparison to related multistate-capable methods
- No head-to-head baselines against tied/dynamic MPNNs, ProteinGenerator, ProDiT, or diffusion models with multi-state conditioning.
- Actionable step: implement matched-budget baselines and report comparative success rates, diversity, and compute.
- Reproducibility and robustness details are sparse
- Seed sensitivity, variance across runs, and diversity/novelty of sequences are not characterized.
- Actionable plan: report per-seed variability, sequence/structure diversity metrics, and failure mode taxonomy.
- Open method question: can SwitchCraft be elevated from per-design optimization to amortized design?
- Current approach optimizes each sequence from scratch; no attempt to train a conditional generator that internalizes multistate constraints to improve efficiency.
- Actionable direction: train amortized models (e.g., diffusion or autoregressive) using SwitchCraft losses as training signals or as RL rewards.
Practical Applications
Immediate Applications
The following applications can be deployed now in silico and used to drive near-term experimental work, leveraging the open-source SwitchCraft code and demonstrated workflows.
- Ligand-responsive fluorescent biosensor prototyping (sectors: healthcare, diagnostics, synthetic biology, education)
- Use case: Rapidly generate small-molecule-responsive conformational switches and fuse them to cpGFP to pre-screen candidate biosensors (e.g., SAM, cGMP, ATP), prioritizing constructs that modulate chromophore contacts upon ligand binding.
- Tools/workflows: SwitchCraft loss templates (BindingLoss, ContactLoss, ConfChangeLoss), automated insertion-site selection into cpGFP, chromophore-contact screening; bench follow-up with gene synthesis, expression, fluorescence assays, and FACS.
- Assumptions/dependencies: Structure predictions (Boltz-1) correlate with function; ligand parameterization and ion/DNA handling are accurate; constructs express and fold; wet-lab validation and optimization (linkers, expression hosts) required.
- Conditional binders and allosteric switches for research tools (sectors: drug discovery, synthetic biology, academic research)
- Use case: Design proteins that bind a peptide/protein only in the presence of an effector ion/small molecule (induced binding) or toggle motif exposure/function via positive/negative allostery.
- Tools/workflows: Induced-binding specifications with state-partitioned BindingLoss/anti-BindingLoss; SPR/yeast display/biolayer interferometry for affinity characterization; cell-based assays for conditional recruitment.
- Assumptions/dependencies: In silico iPTM and contact confidence correlate with binding; proper metal coordination and effector concentrations are achievable in vitro/in vivo; kinetics/pathway feasibility not modeled.
- Multiplex analyte discrimination miniproteins (sectors: diagnostics, environmental monitoring, synthetic biology)
- Use case: Design small proteins adopting distinct conformations for different ligands (ligand discrimination) and couple them to FRET pairs or split reporters for multi-analyte sensing.
- Tools/workflows: Three-state specifications (apo + two ligands) with per-state losses; downstream reporter engineering; plate-based or microfluidic screening of response curves.
- Assumptions/dependencies: Specificity and orthogonality persist outside the prediction model; small proteins remain stable and non-aggregating; cross-reactivity managed through selection/evolution.
- Motif switching to create dual-function research reagents (sectors: academia, enzyme engineering)
- Use case: Toggle between two motifs (e.g., regulatory or catalytic mini-motifs) with a single effector to study causality, control activity windows, or prototype dual-purpose enzymes.
- Tools/workflows: MotifSpec merging, motif RMSD-driven filters, state-specific anti-motif terms; lab validation via activity assays or structural methods (cryo-EM/NMR for larger systems).
- Assumptions/dependencies: Motif placement guided by structural losses reflects true activity; insertion context and loop design tolerate switching without misfolding.
- Community benchmarking and method development (sectors: academia, software)
- Use case: Adopt the paper’s positive/negative allostery battery (11/24 motifs succeeded across diverse ligands) as a standardized multistate benchmark for new design algorithms.
- Tools/workflows: Public codebase, curated motif/ligand sets, evaluation metrics (mRMSD, intraRMSD, crossRMSD, iPTM).
- Assumptions/dependencies: Community consensus on metrics and success thresholds; reproducibility across hardware and model versions.
- Software products and services for multistate protein CAD (sectors: software, biotech R&D)
- Use case: Offer a cloud or on-prem design environment with a DSL for specifying multistate objectives and a library of reusable losses (motif, binding, conformational change).
- Tools/workflows: PyTorch/Boltz-1 pipelines, queueing for multi-state evaluations, result dashboards, LIMS integration.
- Assumptions/dependencies: Access to GPUs; model/checkpoint licensing; robust ligand/DNA/ion input pipelines; user training.
Long-Term Applications
These opportunities require further research, scaling, and/or validation, including kinetics-aware constraints, improved ligand handling, and rigorous wet-lab testing.
- Controllable therapeutics and ligand-gated biologics (sectors: healthcare, pharma)
- Use case: Antibodies, cytokines, and binders that activate only in the presence of disease-specific metabolites/ions; small-molecule-gated CAR-T or cell therapies; prodrug-activating enzymes with safety switches.
- Tools/workflows: Multistate designs with anti-binding in off-states, target-ligand context modeling, cell-based functional screens, toxicology and immunogenicity panels.
- Assumptions/dependencies: Human safety, immunogenicity, PK/PD, and manufacturability; regulatory approvals; kinetics and off-target binding minimized; robust performance in complex biofluids.
- Multi-step de novo enzymes and molecular machines (sectors: enzyme engineering, industrial biotech, fundamental science)
- Use case: Catalysts that progress through designed intermediates with distinct conformations; ligand- or redox-driven rotary/motor-like proteins; GPCR-like multi-ligand logic.
- Tools/workflows: Expanded loss library for transition-state proxies and pathway constraints; integration with QM/MM, MD, or learned energy landscapes; directed evolution for rate optimization.
- Assumptions/dependencies: Accurate modeling of catalytic geometries and kinetics; handling of cofactors, protonation states, and water networks; stability under process conditions.
- Programmable protein logic for synthetic circuits (sectors: synthetic biology, cell engineering)
- Use case: Multi-input/multi-output protein logic gates controlling localization, transcription, or signaling via state-dependent interfaces; orthogonal switches for layered circuits.
- Tools/workflows: Multi-ligand multi-state objective composition; circuit-level modeling; standardized parts for expression, degradation tags, and localization.
- Assumptions/dependencies: Predictable expression and orthogonality in cells; minimal retroactivity and crosstalk; tunable thresholds and hysteresis.
- Smart biosensors for wearables and point-of-care diagnostics (sectors: diagnostics, consumer health, public health)
- Use case: Stable, shelf-ready protein sensors for metabolites, drugs, and pollutants in cell-free or encapsulated formats; real-time monitoring of therapy adherence or exposure.
- Tools/workflows: SwitchCraft-designed switches coupled to ratiometric reporters; lyophilization/formulation; device integration (optics, microfluidics).
- Assumptions/dependencies: Long-term stability, batch consistency, minimal matrix effects; regulatory clearance; large-scale manufacturing.
- Industrial biocatalysts with switchable activity (sectors: chemicals, food, energy)
- Use case: On-demand activation/inactivation to synchronize with feedstock changes; process safety via ion- or small-molecule-triggered shutoff.
- Tools/workflows: Multistate enzymes responsive to process cues; process analytical technology (PAT) feedback; reactor control strategies.
- Assumptions/dependencies: Solvent tolerance, temperature stability, and scalability; low-cost cofactors; regulatory compliance for food/pharma.
- Responsive biomaterials and nanodevices (sectors: materials, robotics at the nanoscale)
- Use case: Protein-based hydrogels or coatings that modulate stiffness, porosity, or binding upon ligand/ion exposure; nanoscale actuators for controlled release.
- Tools/workflows: Crosslinkable switch proteins, material integration protocols, mechanical and diffusion characterization.
- Assumptions/dependencies: Mechanical robustness, cycling stability, biocompatibility; consistent behavior in complex environments.
- Closed-loop autonomous DBTL for multistate proteins (sectors: lab automation, biotech R&D)
- Use case: Iterative propose–build–test–learn cycles where SwitchCraft proposes designs, robotics execute builds/screens, and active learning refines objectives and priors.
- Tools/workflows: High-throughput cloning/expression, multiplex assays (binding, fluorescence, activity), Bayesian optimization and reinforcement learning layers.
- Assumptions/dependencies: Capital investment in automation; standardized data schemas; robust coupling between in silico metrics and experimental readouts.
- Standards, governance, and IP frameworks for AI-designed proteins (sectors: policy, legal, standards bodies)
- Use case: Benchmarks (e.g., allostery suite) as community standards; guidance for disclosure, safety assessment, and environmental release; IP strategies for programmatic design artifacts and specifications.
- Tools/workflows: Consortium-driven datasets and challenge problems; safety-by-design checklists; model cards and validation protocols.
- Assumptions/dependencies: Multi-stakeholder consensus; evolving regulatory landscapes; dual-use risk management.
Notes on feasibility across applications:
- Core dependencies: accuracy and generalization of structure-prediction-guided losses; realistic ligand/ion/DNA handling; kinetic plausibility (currently not enforced); expression, stability, and solubility in target hosts; availability of GPU compute and model checkpoints.
- Practical mitigations: incorporate MD or learned energy/kinetic constraints; expand ligand parameter libraries; integrate negative design for off-target states; rely on directed evolution to refine promising in silico hits.
Glossary
- Allostery: Regulation of a protein’s activity via ligand-induced changes at a site distinct from the functional site; here used for ligand-activated or -deactivated motifs. "single ligands activating (positive allostery) or deactivating (negative allostery) a structural motif"
- Anti-binding loss: A penalty term that discourages protein–ligand binding in specified states. "the anti-binding loss "
- Anti-motif loss: A penalty term that discourages scaffolding of a target motif in specified states. "the anti-motif loss "
- Apo state: The ligand-unbound state of a protein. "a ContactLoss in both apo and holo states"
- Backpropagation through structure prediction models: Using gradients from differentiable structure predictors to optimize sequences toward design objectives. "based on backpropagation through compositional design constraints parameterized by structure prediction models"
- Binding loss: An objective that promotes confident, high-probability contacts between a protein and a ligand. "The binding loss aggregates the confidence of the top two contacts per ligand token for the top tokens"
- Boltz-1: A structure prediction model used as a differentiable oracle in the design loop. "under several instantiations of Boltz-1"
- BoltzDesign-1: A gradient-based protein design protocol whose schedule and losses are adapted in this work. "a multi-stage optimization protocol adapted from BoltzDesign-1"
- Chromophore: The light-absorbing moiety in fluorescent proteins whose environment controls fluorescence. "which quenches fluorescence, while ligand binding induces a conformational change that tugs this residue away from the chromophore"
- Circularly permuted GFP (cpGFP): A GFP variant with new termini near the chromophore, making fluorescence sensitive to local structural changes. "a fluorescent reporter domain such as circularly permuted GFP (cpGFP)"
- Co-folding: Simultaneous structure prediction of a designed sequence with its molecular context(s) to evaluate state-specific losses. "co-folded with each state's molecular context"
- Conformational change loss: An objective that encourages structural differences between specified states. "The conformational change loss maximizes this change for the contact with the largest sensitivity for each residue"
- Conformation switcher: A designed protein module that changes structure upon ligand binding to transmit a signal. "we use SwitchCraft to design conformation switchers that are responsive to three small molecule ligands"
- Distogram: A predicted discrete probability distribution over inter-residue distances output by structure models. "the distogram output of Boltz-1"
- Effector: A molecule (often a ligand) that triggers switching between protein functional states. "a ligand effector"
- Gumbel-softmax: A differentiable sampling/relaxation technique used to initialize or optimize discrete sequence choices. "initializes from a Gumbel-softmax distribution"
- iPTM: AlphaFold’s interface predicted TM-score metric used as a proxy for binding confidence. "effector iPTM "
- IntraRMSD: The average pairwise RMSD among predicted structures within a state, used to assess prediction consistency. "IntraRMSD measures structural consistency by averaging pairwise RMSDs among predicted structures within the same state"
- Jensen–Shannon divergence (JSD): A symmetrized divergence between probability distributions used here to compare distance distributions across states. "and JSD is the Jensen-Shannon divergence"
- Ligand discrimination: Designing distinct protein conformations depending on which of multiple ligands is bound. "that switch between three distinct conformations depending on which of two ligands is bound (ligand discrimination)"
- Ligand modification: Designing proteins whose structure/function changes upon a chemical change in a bound ligand. "that exhibit a large conformational shift upon change to a bound ligand (ligand modification)"
- Motif RMSD (mRMSD): The RMSD between a designed motif region and its target conformation, measuring scaffold accuracy. "Motif RMSD (mRMSD) measures the alignment error of scaffolded motifs relative to their target conformations"
- Motif scaffolding: Embedding a predefined structural motif within a designed protein backbone. "the well-known RFDiffusion motif scaffolding benchmark"
- pLDDT: Per-residue predicted confidence score from AlphaFold-like models, used to filter reliable designs. "average pLDDT "
- pTM: Predicted TM-score from AlphaFold-like models indicating global structure confidence. "pTM=0.60"
- Radius of gyration: A measure of structural compactness used to filter designs. "radius of gyration $\leq 22 \si{\angstrom}$"
- RFDiffusion: A diffusion-based protein structure generator often used for motif scaffolding and binder design. "the well-known RFDiffusion motif scaffolding benchmark"
- Straight-through estimator (STE): A technique that enables gradient flow through discrete argmax operations via a surrogate. "a straight-through-estimator (STE) is utilized"
- State switching: The ability of a protein to adopt multiple functional conformations depending on context or ligands. "designing state-switching proteins"
Collections
Sign up for free to add this paper to one or more collections.