- The paper introduces a novel application of q-statistics to model non-Gaussian amplitude fluctuations in EEG signals from both typical and ADHD children.
- It establishes an affine scaling law between the q index and beta parameters across EEG channels, indicating universal critical behavior in brain dynamics.
- Findings reveal that elevated q values in ADHD subjects may serve as potential biomarkers for refining neuropsychiatric diagnostics.
Nonadditive Entropic Analysis of EEGs: Probing Brain Criticality in Typical and ADHD Children
Introduction and Motivation
This work investigates the critical dynamics underlying human brain activity through the lens of nonextensive statistical mechanics, with a specific focus on the electroencephalogram (EEG) signals of boys diagnosed as either typical or with Attention Deficit Hyperactivity Disorder (ADHD). The central motivation stems from the absence of objective, quantitative biomarkers for neuropsychiatric disorders and the increasing recognition that neural dynamics reside at or near critical points, characterized by complex, scale-invariant behavior. Traditional measures fail to capture the heavy-tailed, non-Gaussian features seen in neural signals, prompting the need for generalized statistical descriptions.
Methods: q-Statistics for Non-Gaussian Signal Structure
EEG signals were recorded from 19 typical and 19 ADHD boys during an Attention Network Test. Standard preprocessing was applied, including band-pass filtering, power-line artifact removal, zero-meaning, and clipping of extreme values. Signals were then divided into stationary segments; amplitude fluctuations within each formed the basis of subsequent analysis.
The probability distributions of these normalized amplitude fluctuations were empirically estimated and fitted with q-Gaussian distributions, a family of distributions that generalizes the standard Gaussian via the Tsallis nonadditive entropy framework:
ρ(x)=ρ0eq−βx2,eqx=[1+(1−q)x]+1/(1−q)
where q is the entropic index (with q=1 recovering the Gaussian), β is the scale parameter, and ρ0 normalizes the distribution.
The fitting approach utilized a transformation y=lnq(ρ/ρ0) to linearize the fitting in terms of x2, optimizing q to maximize the coefficient of determination q0. Values of q1 and q2 were extracted for each channel, participant, and segment, supporting detailed statistical comparisons between the typical and ADHD groups.
Figure 1: Signal segments for the Fz and O1 channels for four arbitrarily chosen patients, highlighting the pronounced amplitude fluctuations.
Results: Emergent q3-Gaussian Structure and Scaling
Analysis demonstrated that EEG amplitude fluctuations in both cohorts consistently deviate from Gaussian statistics, instead showing excellent fit to q4-Gaussians with q5. This confirms the presence of nonlocal correlations and heavy tails in the neural signal distribution (Figure 2):
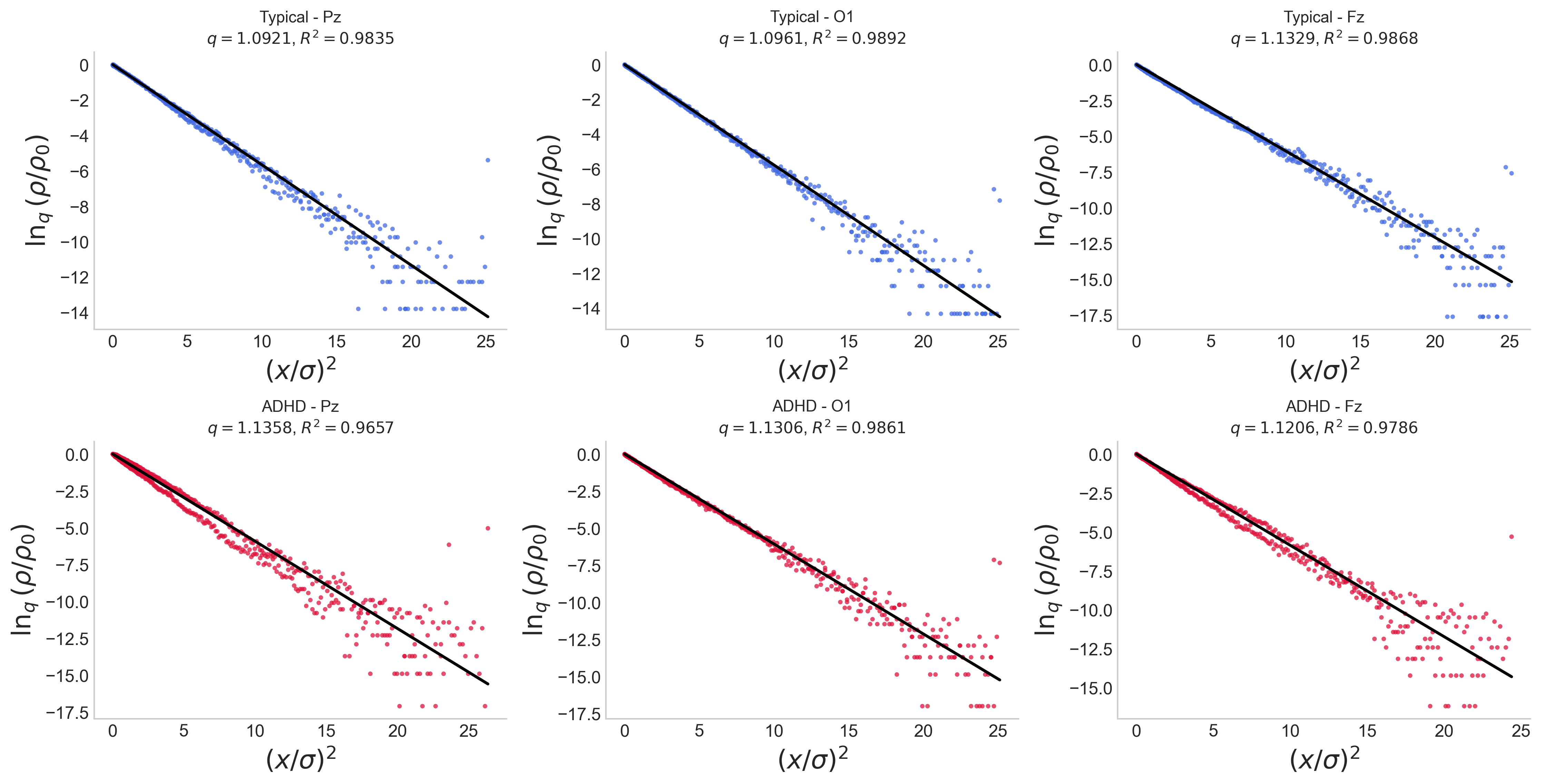
Figure 2: Linear relationship between q6 and q7 for selected channels, illustrating compatibility with the q8-Gaussian hypothesis.
A key finding is the empirical scaling law relating the parameters q9 and ρ(x)=ρ0eq−βx2,eqx=[1+(1−q)x]+1/(1−q)0. Across patients and EEG channels, ρ(x)=ρ0eq−βx2,eqx=[1+(1−q)x]+1/(1−q)1 was found to depend affinely on ρ(x)=ρ0eq−βx2,eqx=[1+(1−q)x]+1/(1−q)2:
ρ(x)=ρ0eq−βx2,eqx=[1+(1−q)x]+1/(1−q)3
with exponents ρ(x)=ρ0eq−βx2,eqx=[1+(1−q)x]+1/(1−q)4 and intercepts ρ(x)=ρ0eq−βx2,eqx=[1+(1−q)x]+1/(1−q)5 differing weakly across channels. This scaling persisted in both ADHD and typical groups.
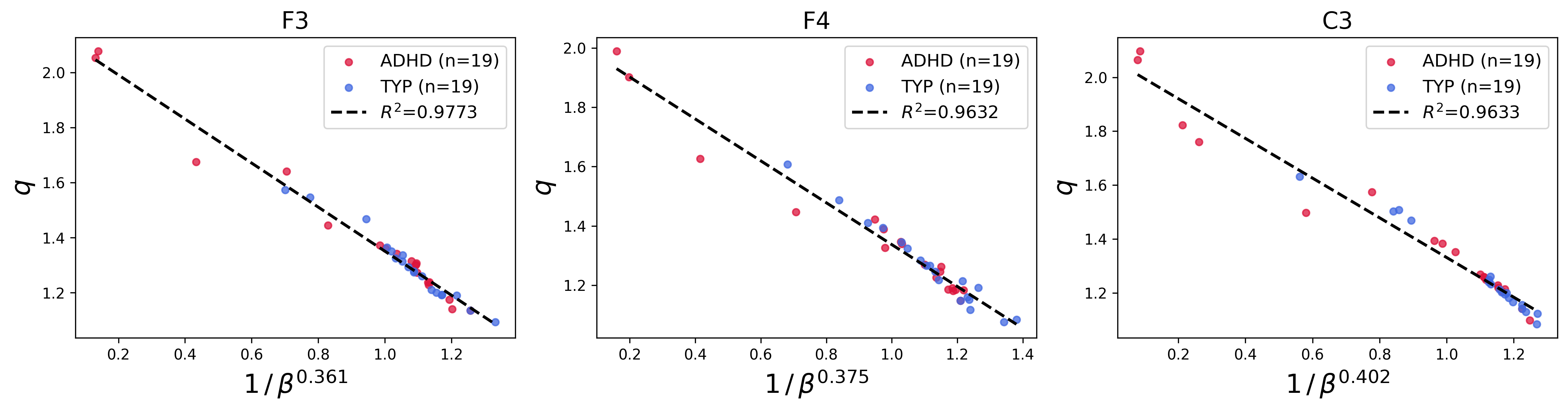
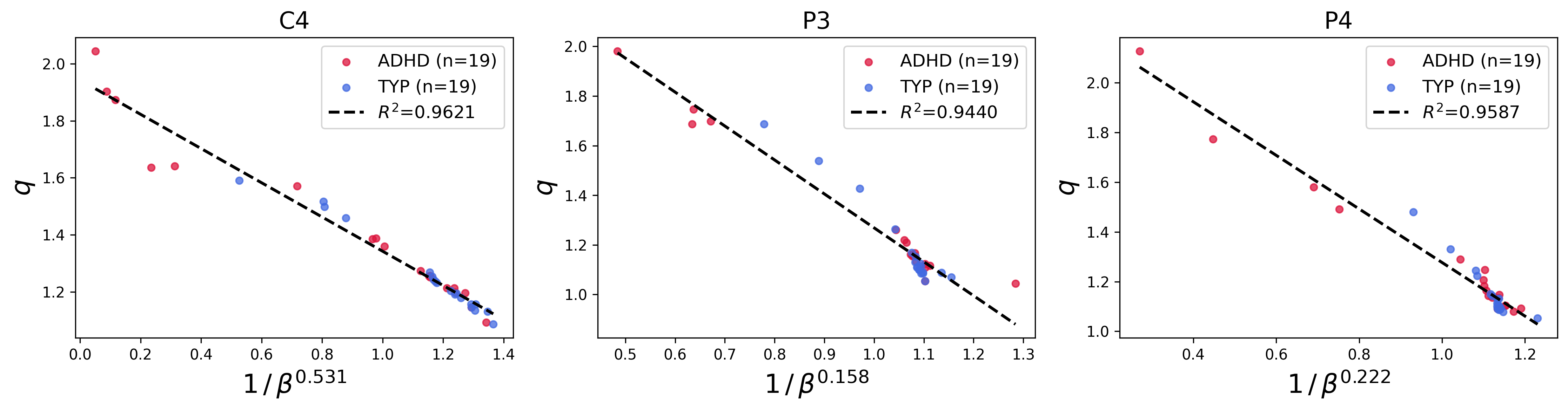
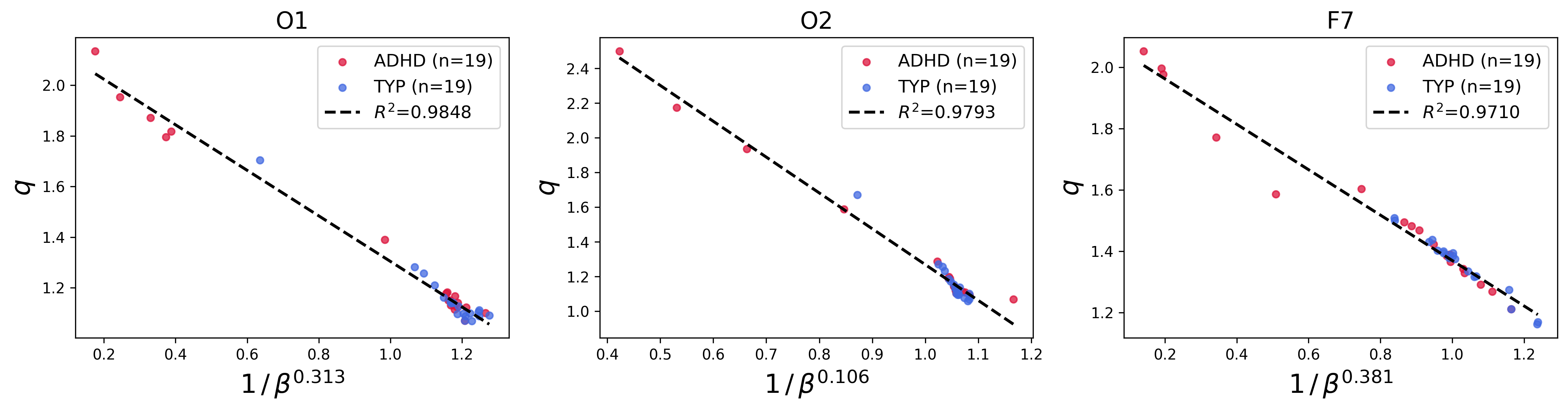
Figure 3: Channel-specific plots of ρ(x)=ρ0eq−βx2,eqx=[1+(1−q)x]+1/(1−q)6 versus ρ(x)=ρ0eq−βx2,eqx=[1+(1−q)x]+1/(1−q)7 for all patients, demonstrating the affine scaling law across multiple EEG sensor locations.
Global analysis across all channels and all subjects revealed a striking data collapse, implicating a shared universality class for brain criticality in both ADHD and typical populations (Figure 4):
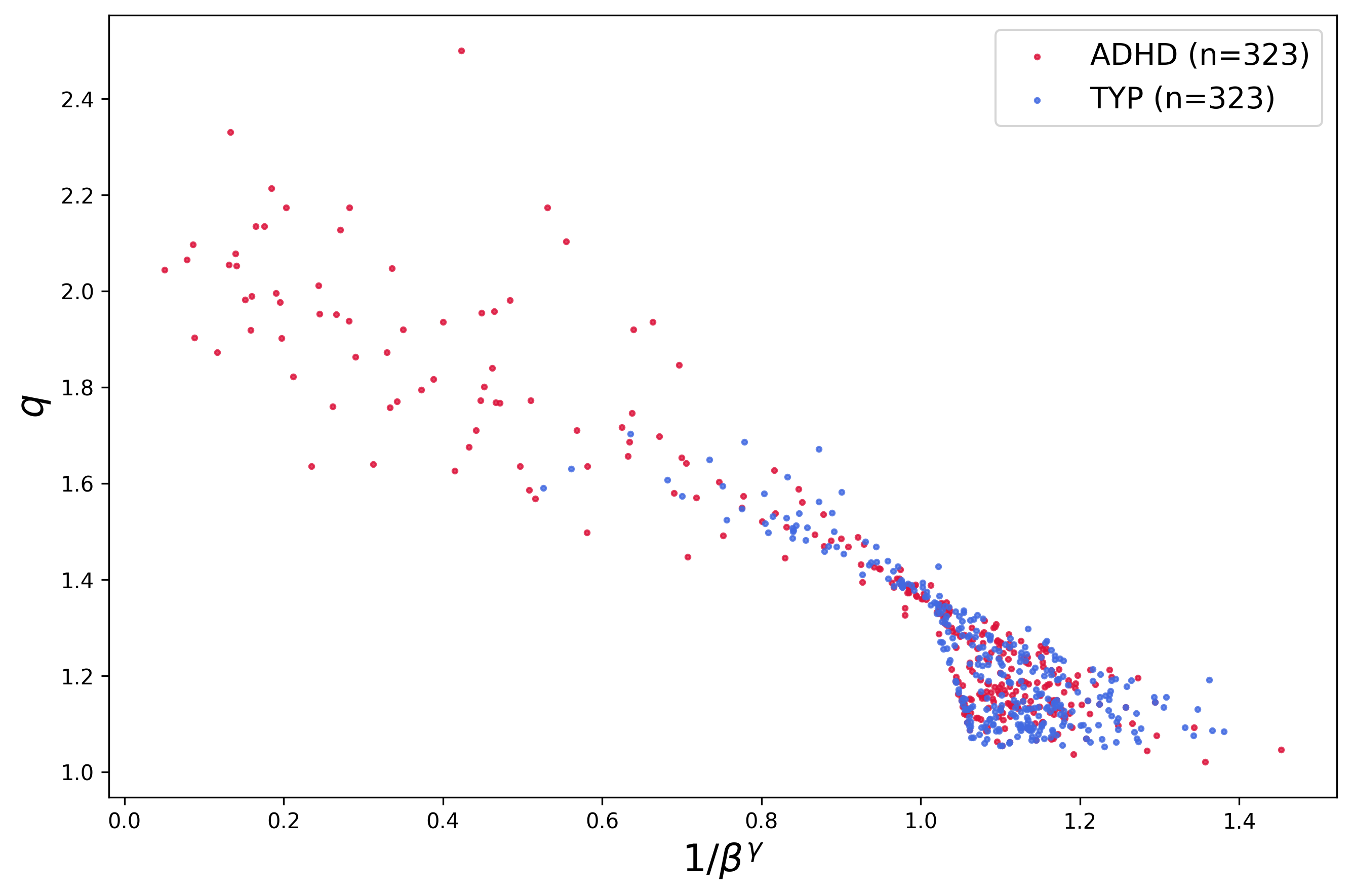
Figure 4: Plot of ρ(x)=ρ0eq−βx2,eqx=[1+(1−q)x]+1/(1−q)8 versus ρ(x)=ρ0eq−βx2,eqx=[1+(1−q)x]+1/(1−q)9 across all 17 channels and 38 patients, reinforcing the universality of the critical scaling.
Yet, detailed inspection revealed that ADHD subjects tended to have higher mean q0 values and increased inter-individual variability in q1 (and correspondingly in q2), implying a segregation in complexity, with ADHD brains showing higher statistical complexity.
Channel-wise breakdown revealed the absence of strong spatial anomalies for typical children. In contrast, a subset of ADHD participants exhibited pronouncedly elevated q3 values for specific channels, potentially marking distinct complexity regimes:
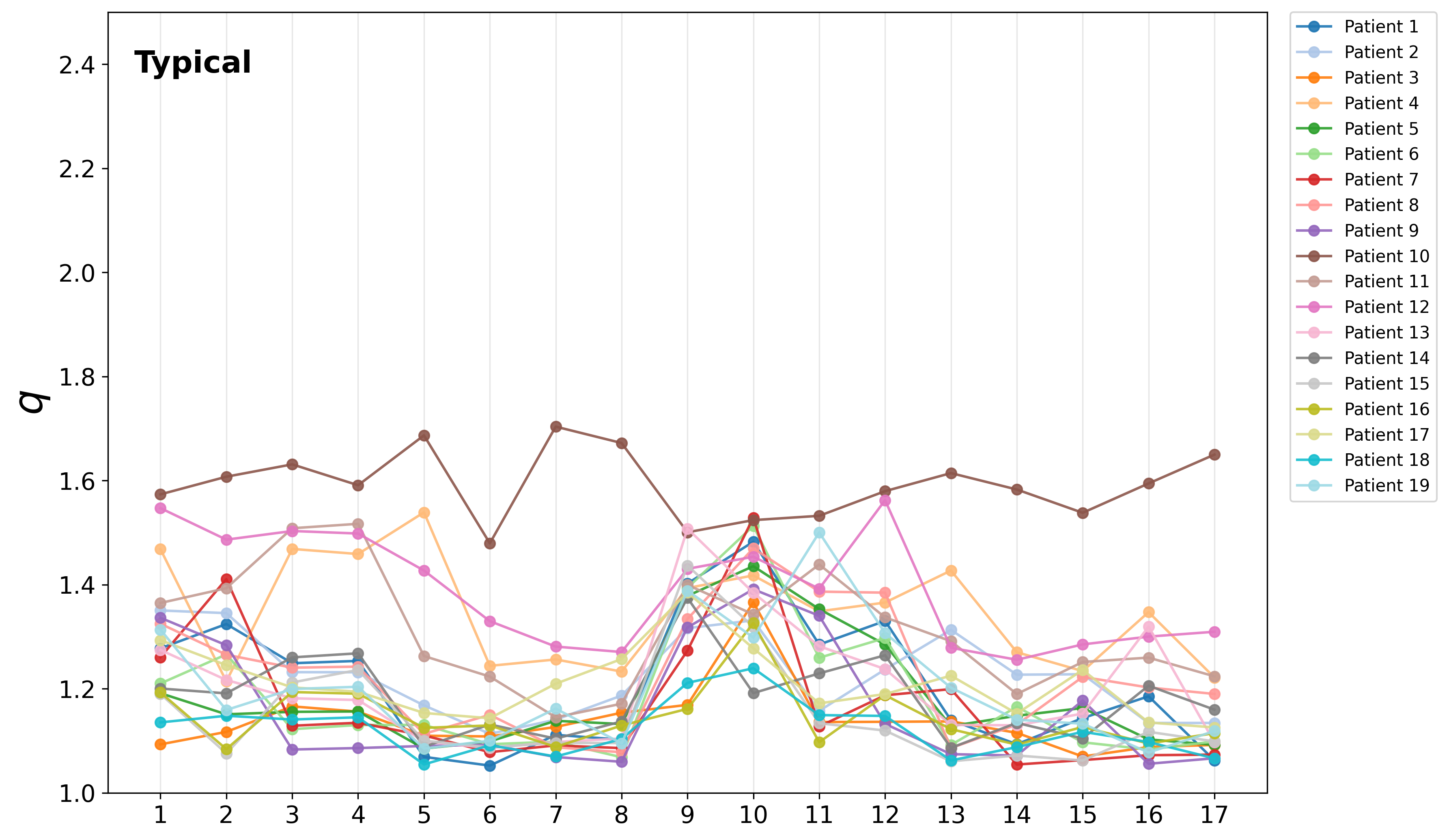
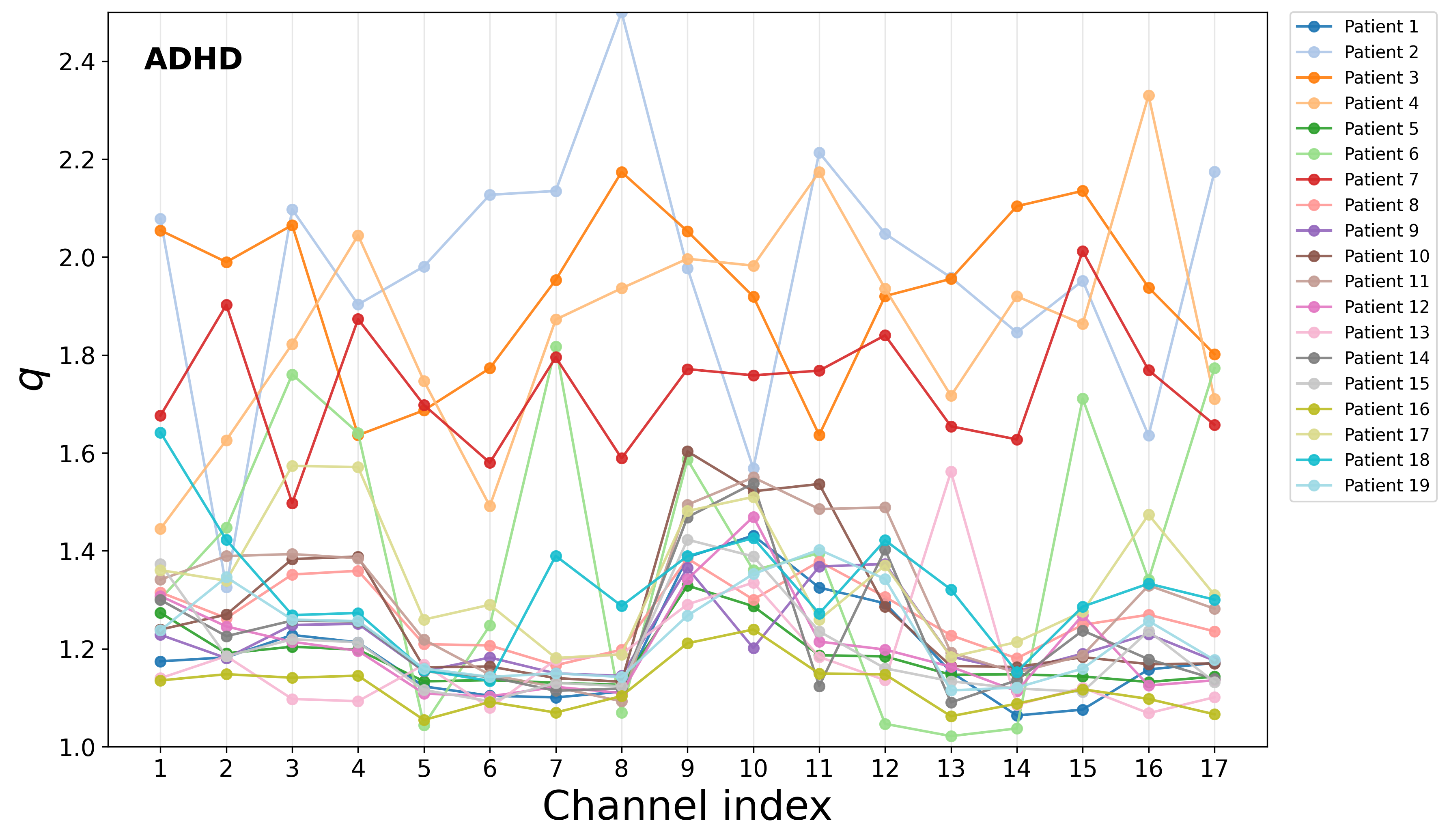
Figure 5: Channel-wise q4 indices for all participants, marking individuals with distinctly elevated complexity parameters mainly in the ADHD group.
Averaging q5 over all channels and ordering participants revealed discrete jumps corresponding to some ADHD-diagnosed patients, potentially suggesting limitations in current categorical diagnoses or the existence of subpopulations with pronounced neurodynamic irregularities (Figure 6):
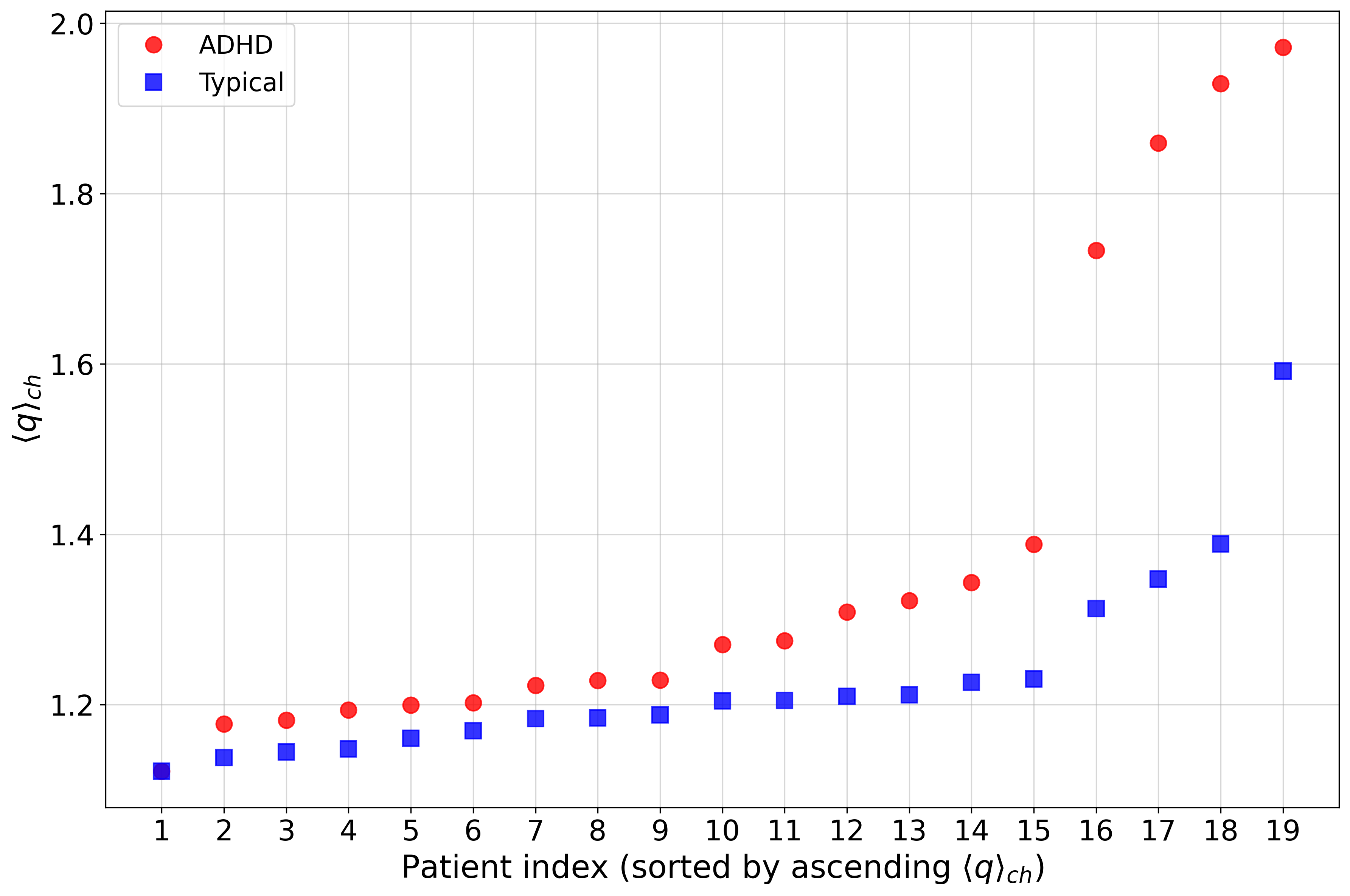
Figure 6: Channel-averaged q6 as a function of patient index, highlighting outlier subjects—especially among the ADHD cohort.
Discussion and Implications
The results robustly indicate that EEG amplitude fluctuations in both typical and ADHD subjects are governed by non-Gaussian, q7-generalized statistics. The q8 index serves as a sensitive metric of neural complexity, demonstrating both group-level shifts and individual variability. Crucially, the parameter scaling law (q9 vs. q=10) suggests that brain dynamics in both cohorts inhabit a critical regime characterized by universality, consistent with self-organized criticality observed in diverse complex systems.
These findings have several implications:
- Objective Biomarker Potential: The q=11-index, and possibly its channel-averaged variant, holds promise as a quantitative biomarker for ADHD and, more generally, for pathologies marked by altered neural complexity. However, significant overlap between groups and the presence of outliers caution against simplistic diagnostic thresholds.
- Critical Dynamics and Universality: Both typical and ADHD brains exhibit dynamics near criticality, as supported by the observed data collapse and scaling laws. This universality parallels theoretical results in condensed matter and nonequilibrium statistical physics, highlighting the deep analogies between brain activity and critical phenomena.
- False Positives and Diagnostic Challenges: The indistinguishability in many cases, combined with distinct outliers, raises the question of false positives or different ADHD subtypes, urging refinement and triangulation of diagnostic practices.
- Gender and Broader Application: The analysis, confined to boys, anticipates studies in girls for whom ADHD manifestations and etiology remain controversial. Extending such complexity analysis across genders and other neuropsychiatric conditions could clarify both biological and behavioral underpinnings.
Conclusion
This study provides compelling evidence that both typical and ADHD neural activities in children, as indexed by EEG amplitude fluctuations, are well described by q=12-Gaussian statistics, confirming the non-Gaussian and complex nature of neural dynamics. The observed affine relation between q=13 and q=14 is a signature of critical phenomena and universality across both diagnostic groups. While q=15 serves as a theoretically motivated complexity measure with diagnostic potential, its overlap between populations and sensitivity to inter-individual variability signal the need for larger studies and continuous refinement. This nonadditive entropic framework offers a robust foundation for quantifying brain criticality, advancing both the physics of complex systems and the search for objective metrics in clinical neuroscience.

