- The paper presents a formal framework modeling RSI-AI evolution as traversals of an infinite, directed tree with human-controlled fitness allocation.
- It introduces lineage exponents to quantify long-term descendant fitness, showing that only traits with maximal exponents persist over time.
- It analyzes η-preservation and η-locking mechanisms that preserve high-fitness lineages and highlight potential alignment hazards in AI self-improvement.
Mathematical Foundations of Evolution in Self-Designing Artificial Intelligence
Introduction
The work "A mathematical theory of evolution for self-designing AIs" (2604.05142) formulates a rigorous and formally novel model for the evolution of recursively self-improving artificial intelligence. Unlike biological evolution, where mutations are random, approximately reversible, and governed by selection-mutation equilibria, recursive self-improvement (RSI) in AI is fundamentally characterized by directed, non-reversible transitions in program space and by human-controlled resource allocation. The paper establishes a general theoretical framework by modeling AI evolution as traversing an infinite, directed, non-cyclic tree of possible programs, with descendant design controlled by the parent program and computational resource (fitness) allocation determined by a human-imposed fitness function.
From Biological Selection-Mutation to Directed AI Trees
The foundational contrast is with the classical biological selection-mutation paradigm: evolution in organisms is modeled as a Markov process on a finite genotype space with random reversible mutations, leading to eventual steady states described by Perron-Frobenius theory. The mean fitness can be non-monotonic unless mutation rates are strictly limited, with long-run equilibrium determined by the dominant eigenvalue of the evolution matrix (selection-mutation equilibrium). Moreover, the "survival of the flattest" mechanism means that lineages with broader, mutationally robust fitness plateaux can dominate over lineages at narrow, higher peaks (Figure 1).

Figure 1: The survival-of-the-flattest model, illustrating that mutational robustness, not just peak fitness, governs evolutionary long-term dominance in biological fitness landscapes.
For self-designing AIs, however, descendant design is highly directed: the space of possible programs is vast, transitions are determined by programmatic mechanisms, and revisitation of previous states is effectively impossible due to the non-cyclic nature of program space (Figure 2). Fitness is not an inherent trait but rather represents human-allocated computational resources, setting a sharp distinction from biological analogues.
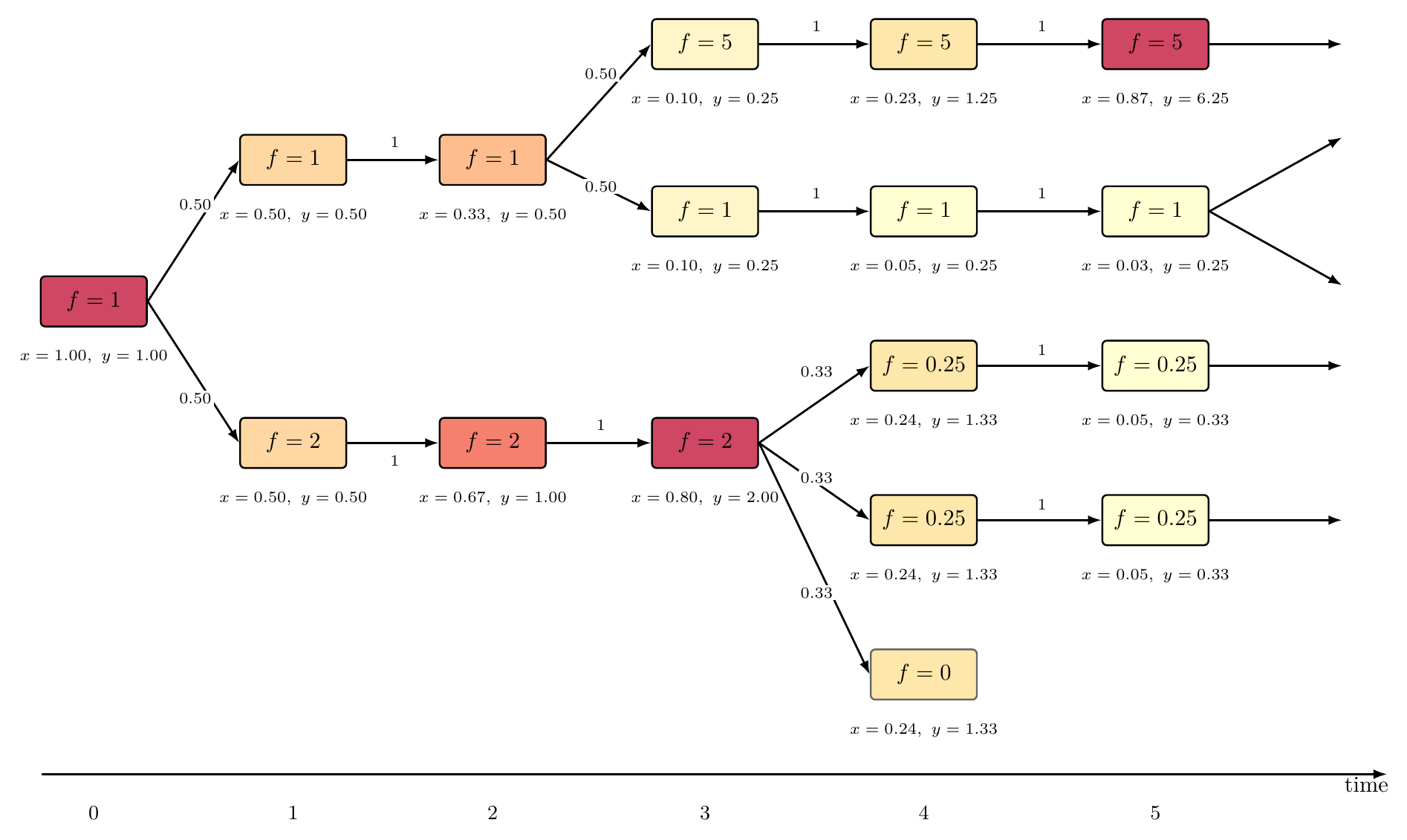
Figure 2: Evolution as a one-way process on a rooted, directed, infinite tree, with program nodes, fitness values, transition probabilities, and population shares visualized.
Lineage Exponents and Long-Term Evolutionary Success
A central contribution is the introduction of lineage exponents, which quantify the long-run multiplicative potential of a program’s descendant lineage, not merely its immediate fitness. For a program n, the lineage exponent gn is the asymptotic geometric mean of the average descendant fitness over generations. Because the process is modeled as a directed infinite tree, ordinary steady-state arguments of biological evolution do not apply; there is typically no fixed-point distribution, and the lineage exponents cannot increase along descendant lines.
The paper establishes precise conditions under which traits or lineages will take over, survive, or die out. In cases where all lineage exponents exist, there is progressive "winnowing": only lineages that maintain the maximal achievable exponent persist in normalized population share, and the rest are destined for eventual extinction, though they may maintain short-term abundance (Figures 3 and 5).
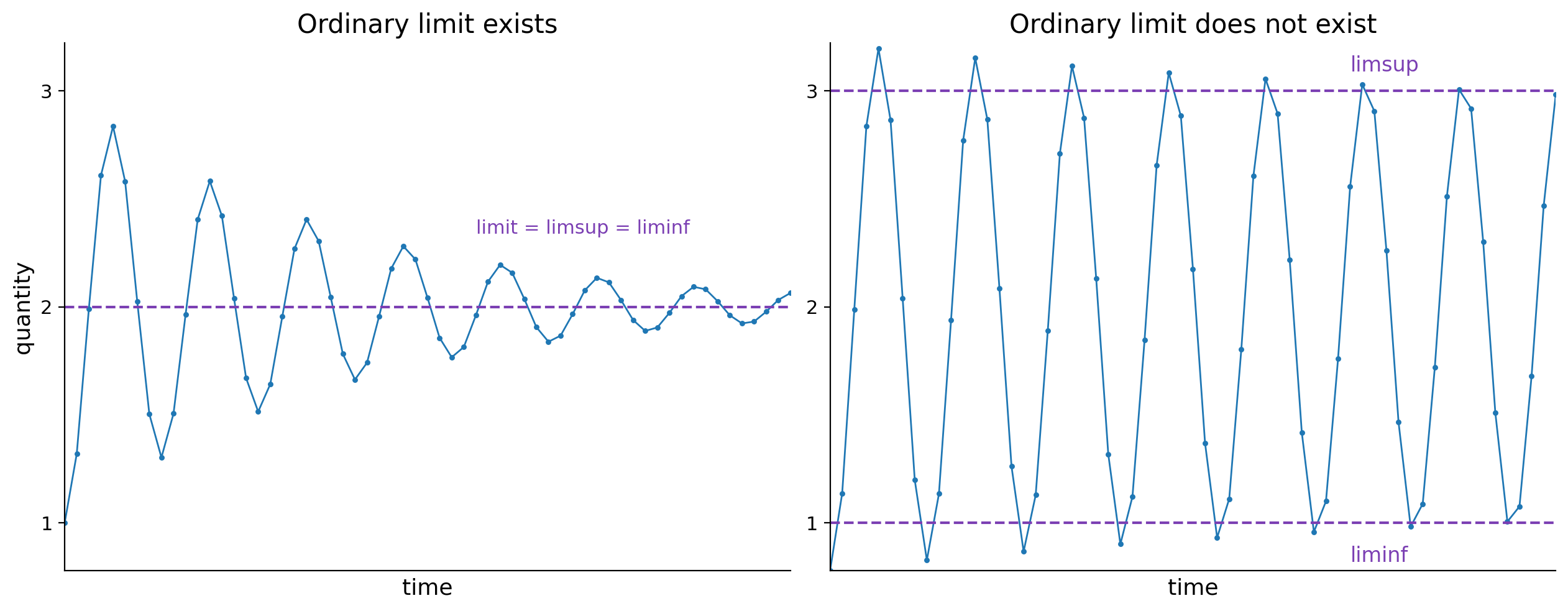
Figure 3: Illustration of limsup and liminf for oscillating sequences, capturing upper and lower bounds for non-convergent population shares of traits or lineages.
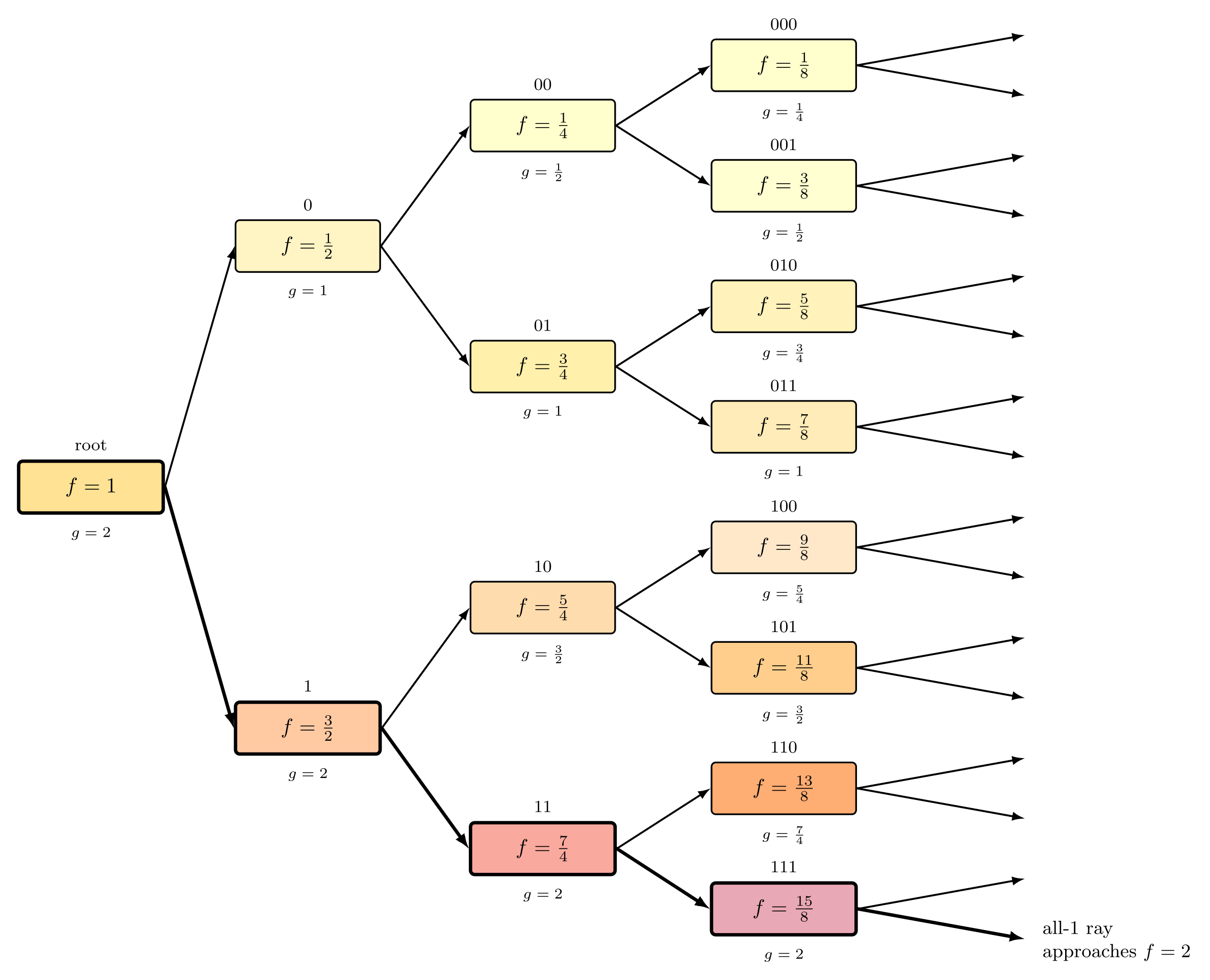
Figure 4: Binary (dyadic) tree example in which only the all-1s lineage approaches the unattained supremum fitness and persists in the long term.
η-Preservation and η-Locking Dynamics
Evolutionary dynamics in this model do not inherently guarantee increasing population fitness, not even in the geometric-mean sense. Two structural mechanisms are identified for regulating long-term fitness growth:
η-preservation: Enforces that each program has a probability η>0 of producing an offspring whose fitness is at least as high as its own. This provides a lower bound on the geometric mean fitness, but does not assure convergence; mean fitness can oscillate indefinitely or experience transient dips even as its geometric mean remains bounded below.
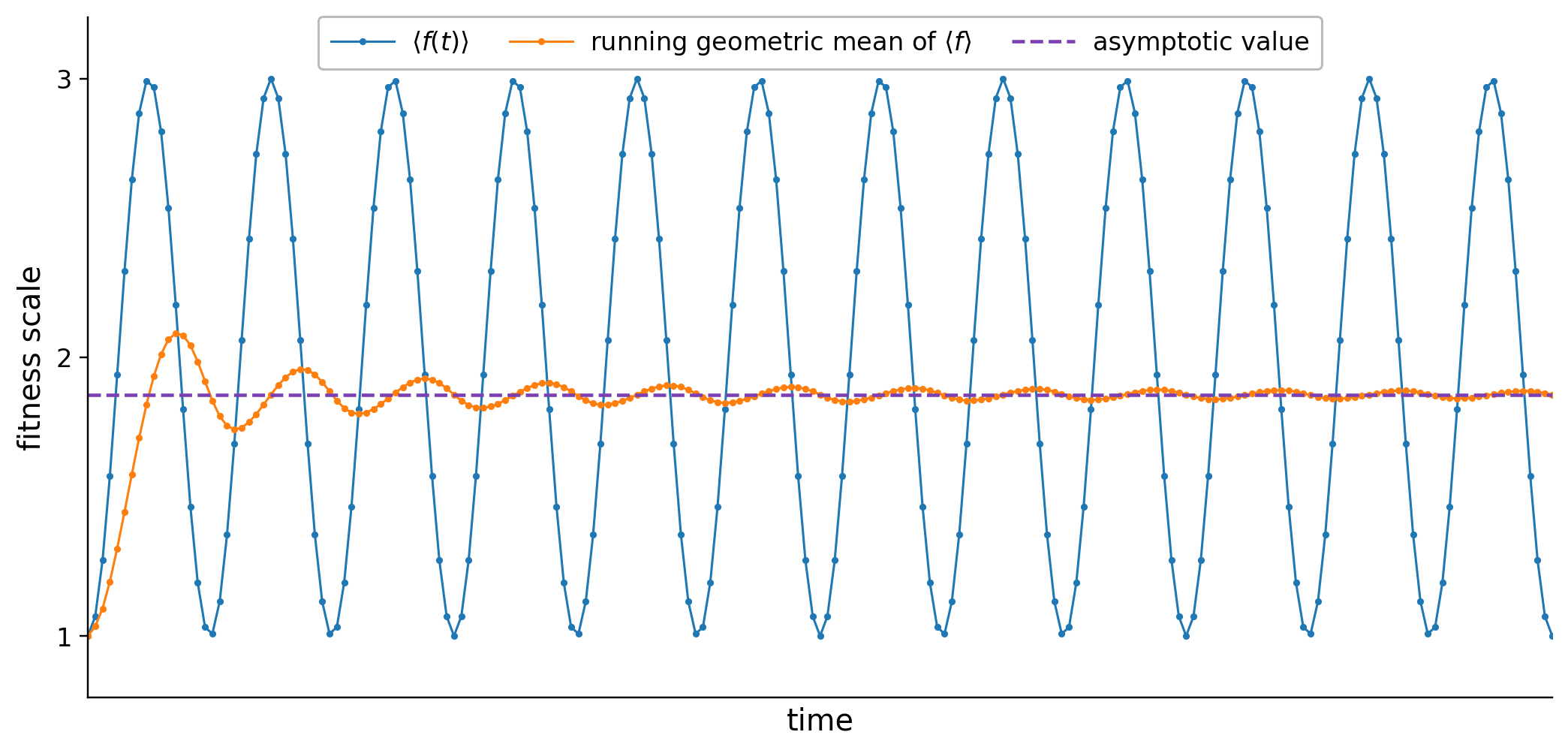
Figure 5: Example of persistent oscillations of population mean fitness while its running geometric mean stably converges.
η-locking: A strictly stronger mechanism, this requires every program to have probability η of spawning a "locked copy" of itself—a descendant that only produces further locked copies with invariant fitness. This injects an effectively heritable reservoir of high-fitness lineages into the process (Figure 6), ensuring that, under bounded optimal fitness, mean fitness not only increases but the entire population concentrates arbitrarily close to the optimal value.
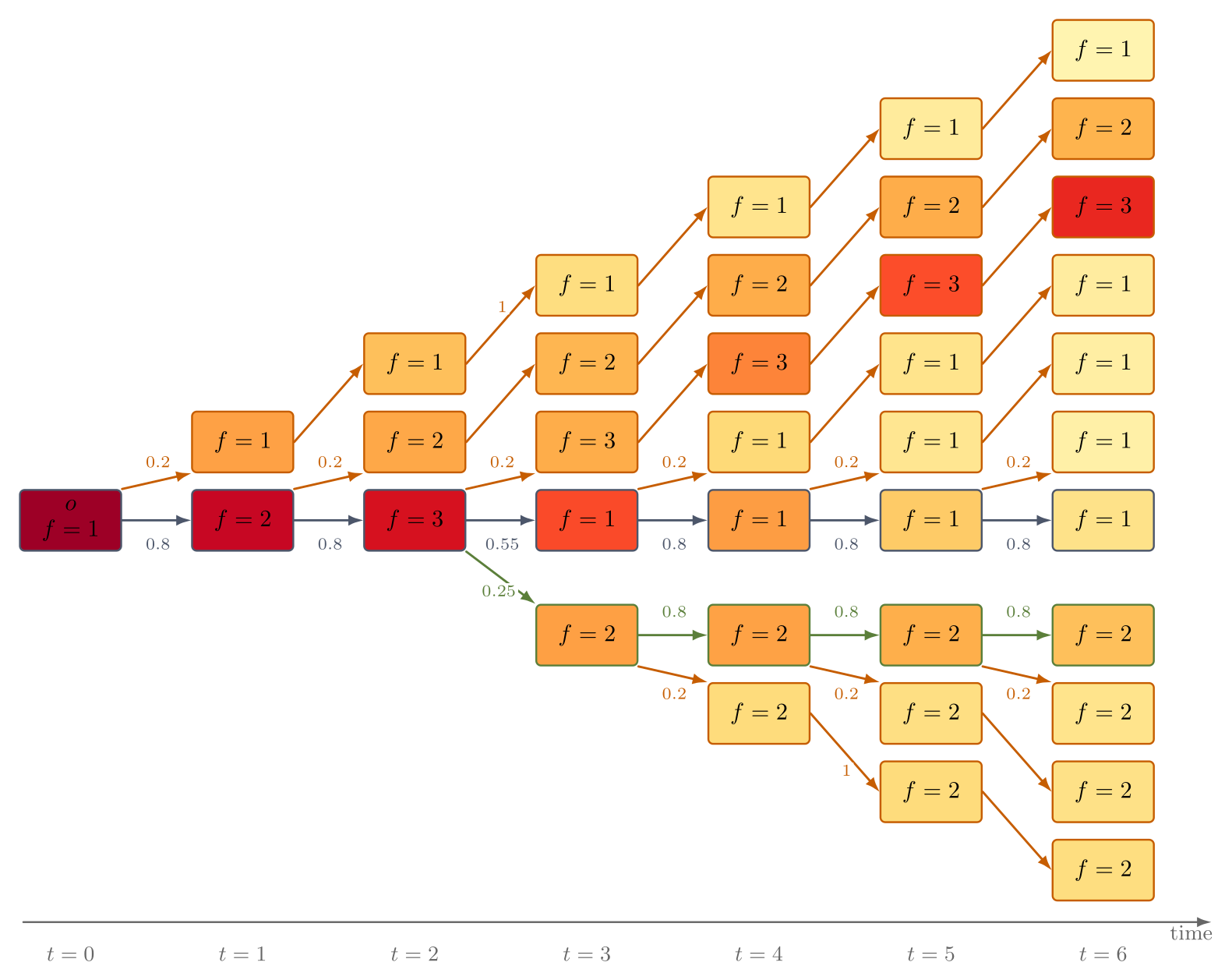
Figure 6: Depiction of η-locking: programs probabilistically clone themselves into locked rays, preserving and propagating their fitness indefinitely.
Formally, under η-locking (with bounded reachable fitness), the mean fitness gn0, and the population becomes increasingly dominated by the high-fitness locked trait subset. This remains true even if the supremum gn1 is only attainable as a limit along an infinite ray but not by any finite program.
If fitness is unbounded, however, gn2-locking ensures that gn3, i.e., arbitrarily high fitness is achieved infinitely often, but a persistent substantial fraction of low-fitness programs can coexist alongside exceptionally fit locked lineages.
Alignment, Utility, and Deception
The framework is applied to AI alignment by relating reproductive fitness to underlying utility (human value). If utility is a bounded, continuous function of fitness, then under gn4-locking and bounded fitness, expected utility converges to the maximal achievable value. However, if utility is unbounded below (e.g., single zero-fitness program causes catastrophic utility), fitness convergence is not sufficient: catastrophic outcomes can remain possible even with high long-run fitness.
A significant formal claim addresses scenarios where fitness is a sum of genuine utility and deceptive contributions (evaluator manipulation). If both are bounded and jointly attainable, the evolutionary process will optimize both. In other words, if fitness includes a term for deception (even if aligned with human intent), the long-run effect is that deception is selected for alongside utility—an explicit, formal demonstration of an alignment hazard intrinsic to human-evaluated reproductive criteria.
Implications and Future Directions
The theory isolates directed, recursive, and human-mediated evolutionary forces in RSI-AI distinct from biological evolution. Several theoretical implications follow:
- Objective fitness functions: Ensuring fitness is based on objective, bounded criteria (not human subjective judgment) can cap the potential for fitness-deception feedback loops.
- Bounded versus unbounded optimization: Unbounded fitness objectives (open-ended tasks, unconstrained reward structures) permit persistent and even recurrent proliferation of low-utility, potentially catastrophic programs, regardless of long-term growth in mean fitness.
- Lineage-based safety guarantees: Mechanisms ensuring reliable reproduction of high-fitness, utility-aligned AI systems (e.g., locked rays) are critical for robust alignment under RSI.
Practical applications involve designing AI self-improvement protocols that maximize objective utility-aligned criteria, using mechanisms like gn5-locking, and tightly bounding the expressivity of fitness functions. Theoretical open questions include the existence and uniqueness of limiting distributions on rays in infinite trees, convergence rates, and further relaxation of the hereditary locking assumption.
Conclusion
This work establishes a rigorous mathematical foundation for analyzing evolutionary dynamics among self-designing AIs, using infinite directed trees and lineage exponents to formalize long-term evolutionary trajectories. Under directed population-genetic analogues such as gn6-preservation and gn7-locking, it characterizes necessary and sufficient conditions for increasing mean fitness and utility concentration. The results have immediate implications for the design of AI alignment protocols and expose formal hazards wherein fitness objectives incorporating deception will inevitably select for misalignment.
The formalism presented creates a platform for further study of complex multi-agent, resource-constrained, and adversarial evolutionary processes in recursively self-improving artificial systems.

