- The paper demonstrates that apparent advantages of connectome topology vanish after rigorous control, revealing initialization and null-model confounds.
- It employs a control ladder methodology to compare naive versus degree-preserving null models in experiments on the Drosophila visual circuit.
- Findings indicate that early performance benefits are likely driven by implementation artifacts rather than intrinsic biological topology.
Topological Sensitivity in Connectome-Constrained Neural Networks: An Analytical Essay
Introduction
This paper, "Topological Sensitivity in Connectome-Constrained Neural Networks" (2604.04033), systematically interrogates the common assertion that biological graph topology—particularly connectome-derived structure—yields measurable improvements in learning efficiency for sparse neural networks. Through rigorous control protocols, the work demonstrates that previously reported advantages are artifacts of initialization and null-model confounds rather than intrinsic properties of connectome topology. The empirical analysis focuses on the Drosophila visual circuit using the flyvis framework, with a granular comparison against both naive and degree-preserving random graph null models.
Control Ladder and Experimental Protocol
The experimental approach is synthesized through a "control ladder" that incrementally strengthens experimental rigor across three axes: (1) initialization procedures, (2) null-model selection, and (3) ensemble robustness. The original weak control—comparing a connectome-constrained neural network against a naive random graph, both initialized from a connectome-trained checkpoint—reveals strong improvements in early loss, network activity, and runtime for the connectome.
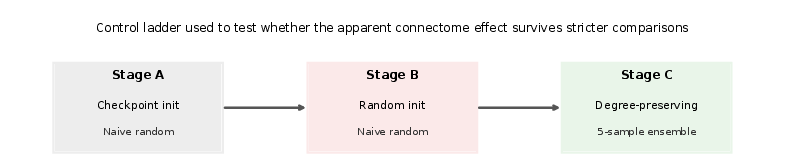
Figure 1: The control ladder delineates the structure of confounds removed through successive experiment stages, culminating in fair from-scratch initialization and degree-preserving null model comparison.
However, when checkpoint initialization is replaced with shared topology-agnostic random initialization, and the naive random null is substituted by a degree-preserving rewired graph (which matches in-degree and out-degree sequences in addition to graph size and self-loops), the apparent connectome advantage vanishes or reverses.
Core Results
The matched-step protocol, focusing on the first 5 and 10 training steps, assesses loss (MSE to direction target), mean absolute network activity, and runtime. Under weak controls (checkpoint, naive random null), the connectome yields a lower loss (0.514 vs. 0.698), reduced mean activity (0.656 vs. 1.861), and faster runtime (252s vs. 309s at 5 steps).
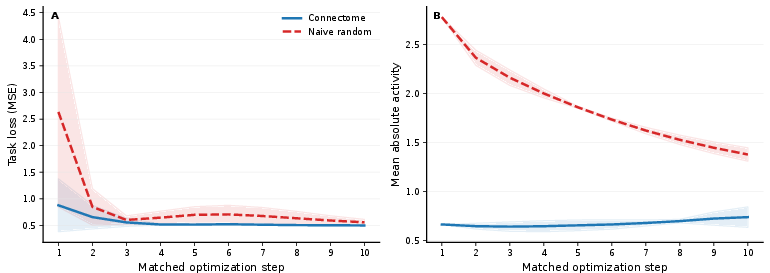
Figure 2: Early training curves under checkpoint and naive null, where the connectome exhibits a strong superficial advantage.
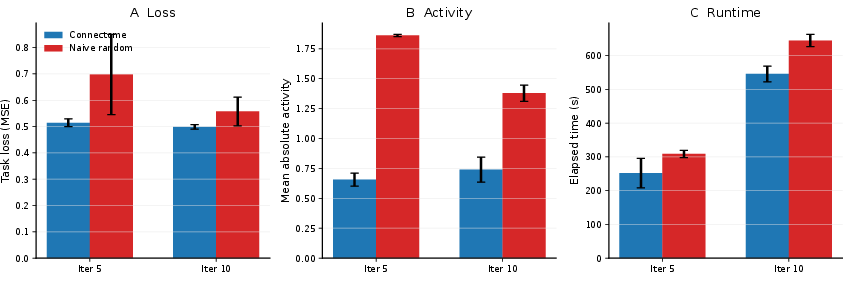
Figure 3: Summary at 5 and 10 steps under weak controls, reaffirming the connectome's superficial outperformance prior to stricter control application.
Once checkpoint bias is removed, the loss advantage disappears (loss difference −0.0020 at both 5 and 10 steps), while slight activity and runtime benefits persist. This residual disappears when the degree-preserving null is introduced: activity differences reverse sign (−0.0106 at 5 steps), and loss remains negligible (+0.0003 at 5 steps). These trends are robust across five independently rewired control graphs, indicating strong ensemble stability.
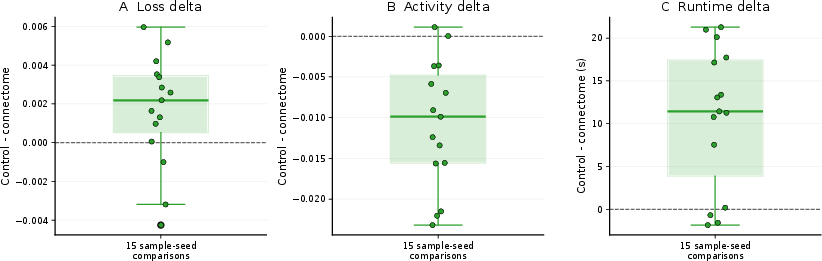
Figure 4: Variability in loss, activity, and runtime across ensemble seed and degree-preserving null samples remains negligible, confirming the robustness of the null.
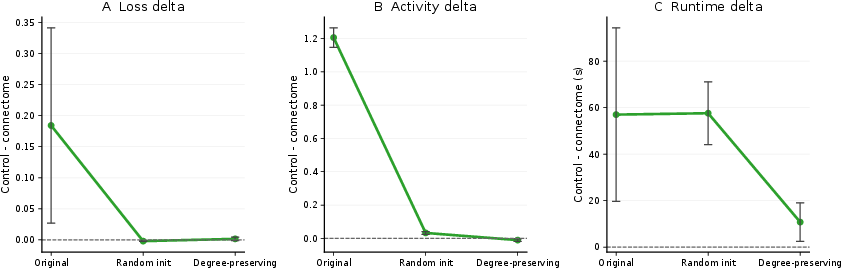
Figure 5: Trajectory of performance delta across control levels—initial loss and activity differences collapse to zero with adequate experimental controls.
Mechanistic Assessment and Metrics
Beyond raw performance, descriptive mechanisms reveal that under weak controls, the connectome structure distributes activity and parameter usage more evenly (lower node and edge Gini coefficients), operates at lower global activity scale, and manifests more distributed routing. Nonetheless, these effects are fully attributable to confounds, as control-overhauled experiments erode all behavioral advantages.
Critically, pre-training diagnostics establish that initialization and gradient scale are matched, excluding trivial numerical mismatches. The runtime advantage for the connectome, though reproducible, is secondary and likely attributable to implementation factors (memory locality, adjacency ordering) rather than graph topology.
Methodological and Theoretical Implications
The findings underscore the methodological primacy of appropriate null-model construction and initialization. Sparse neural network studies must rigorously isolate degree sequence and initialization history as potential confounds; superficial topology effects are otherwise indistinguishable from artifacts of experimental design. This study demonstrates that degree-preserving random rewiring should be standard practice in connectome-based network comparisons.
In the theoretical context, the study delineates the limits of connectome-derived inductive bias in artificial neural networks, especially during early optimization. It calls into question claims of topological optimality when not adequately controlled for degree sequence or prior parameter adaptation.
Limitations
The conclusions are intentionally restricted to the early-training regime, the MovingEdge direction-decoding task, and models structured upon the flyvis parameter-sharing scheme. The analysis does not address long-horizon optimization, convergence properties, alternative architectural or task regimes, or spectral/structural higher-order null models. Further, the study does not disentangle adjacency structure from parameter-sharing correlations inherent in the flyvis metadata tables.
Future Directions
Further exploration is warranted in several areas:
- Scaling to longer training horizons and broader task diversity to determine if topological effects emerge elsewhere.
- Adopting more stringent nulls that match higher-order connectome properties (motifs, clustering, assortativity).
- Explicitly controlling for parameter-sharing and implementation artifacts to fully disentangle topological effects.
- Examining alternate biological connectomes and computational tasks to provide a richer landscape of possible topology-inductive biases.
Conclusion
This work demonstrates that apparent advantages for connectome-constrained neural networks do not robustly persist under fair initialization and degree-preserving null controls, collapsing all performance metrics (loss and activity) to statistical parity. The implication is that observed architectural advantages are confounded by initialization and null-model insufficiency. Only with carefully constructed controls—particularly degree-sequence-matched randomization and rigorous initialization protocols—can the actual effects of biological topology be isolated. This methodological perspective should inform all future work in sparse biological network modeling, connectome-constrained architectures, and structured graph inductive bias studies.