- The paper demonstrates that shot noise dominates ion channel voltage sensing, capping single-channel accuracy at approximately 10 mV.
- It employs a Poisson-Nernst-Planck framework combined with statistical mechanics to model thermal fluctuations and predict sensor geometries.
- The findings indicate that neuronal channel densities are evolutionarily tuned to operate near the physical noise floor established by thermal fluctuations.
Fundamental Physical Limits on Ion Channel Function from Thermal Fluctuations
Introduction and Context
Ion channels are indispensable for neural electrical signaling, acting as discrete voltage sensors embedded in cellular membranes. The operation of these channels, and thus the fidelity of neural signaling, is inherently limited by thermal noise originating from the physical properties of the cellular environment. This work provides a rigorous analysis, grounded in statistical mechanics and electro-diffusive modeling, of the ultimate limits imposed by thermal fluctuations—specifically shot noise and Johnson-Nyquist noise—on the function of voltage-gated ion channels.
The foundational approach parallels, but diverges in critical physical aspects, the classical limits on biological sensing established by Berg and Purcell for diffusive ligand detection. Unlike short-range ligand interactions, ionic communication in the neural milieu is dominated by long-range Coulomb interactions. This hybrid spatial structure of fluctuations leads to distinct noise regimes over biologically realistic length and timescales.
Stochastic Electrostatics of Membrane-Embedded Ion Channels
The dynamics of ions near the neuronal membrane are modeled via the Poisson-Nernst-Planck (PNP) framework, incorporating both diffusive and electric field-driven currents. Introduction of the insulating, low-permittivity lipid bilayer sets up conditions where charge inhomogeneities persist in quasi-two-dimensional surface layers (“Debye layers”), enabling long-range, yet thermally noisy electrical communication. This structure gives rise to two primary categories of noise:
- Shot noise: Fluctuations due to the discreteness of ionic charge, dominant at single-channel and sub-micron spatial scales.
- Johnson-Nyquist noise: Classical voltage fluctuations originating from collective, long-wavelength charge modes, relevant at supra-cellular and ensemble scales.
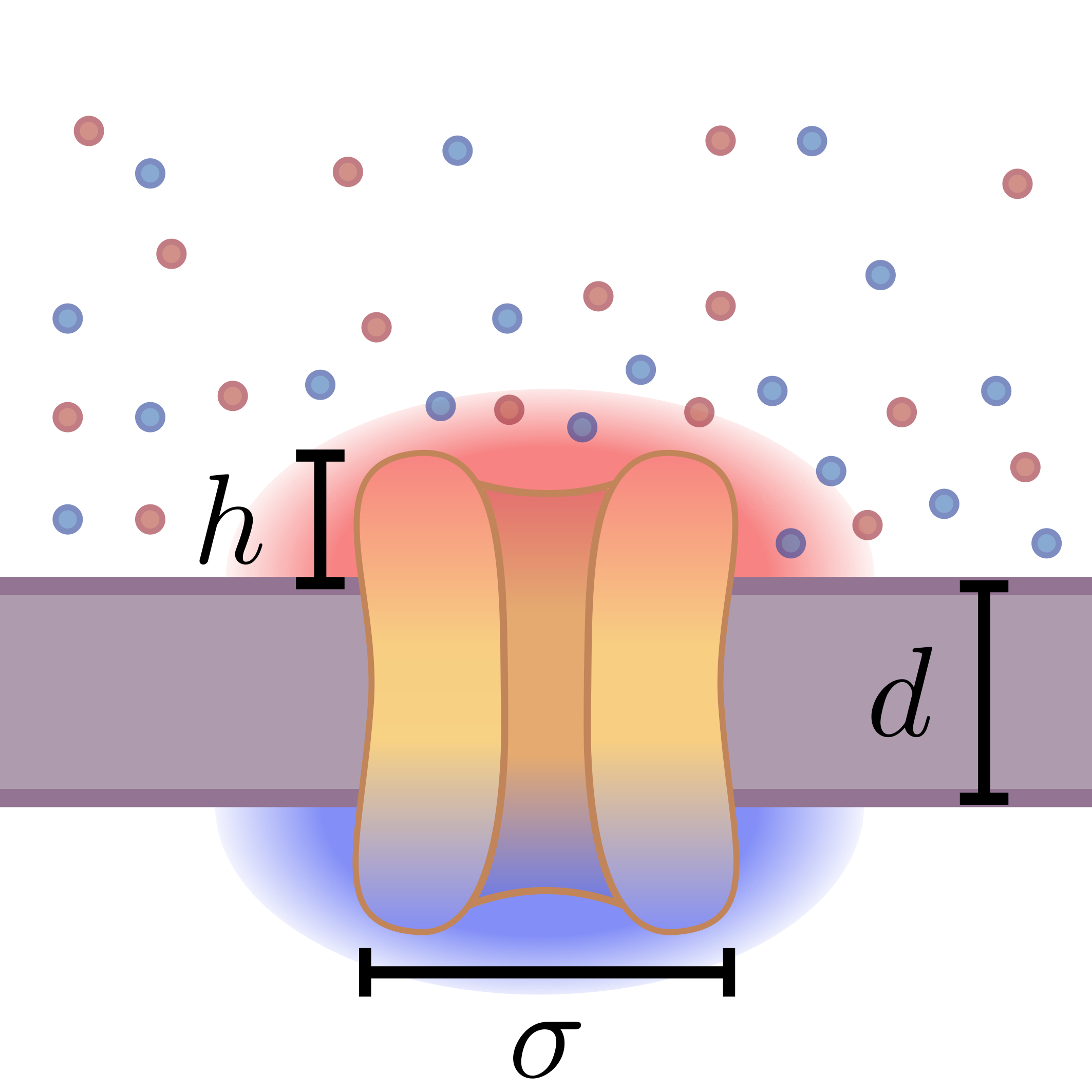
Figure 1: Schematic illustrating the geometric and physical context—an ion channel of width σ and height h, embedded in a membrane of thickness d, sensing the local ionic charge distribution that is fundamentally discrete.
Theoretical analysis, supported by explicit parameter scans, demonstrates that for individual ion channels of physiologically relevant geometry (σ>lD), the noise spectrum is strongly shot noise dominated, with Johnson-Nyquist noise suppressed by low membrane capacitance. The optimal design for voltage sensing is nontrivial, with maximal SNR achieved for channel heights h of order lD, but no significant gain for larger h due to rapid signal saturation.
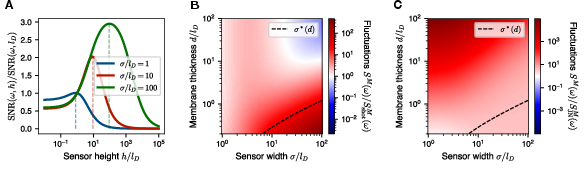
Figure 2: Quantitative comparison of fluctuation amplitudes with analytical predictions for shot noise and Johnson-Nyquist noise, confirming shot noise as the dominant contribution for ion-channel-relevant parameter regimes.
Ultimate Voltage Sensing Precision and Biological Relevance
On the relevant $10$–100μs timescales for biological voltage gating, statistical averaging over the equilibrium shot noise of the sampled ionic charge sets a hard resolution limit. The detection accuracy for a single channel saturates at approximately $10$ mV, within an order of magnitude of empirical gating thresholds and response times measured with single-channel patch clamp setups.
This result implies that voltage-gated sodium and potassium channels operate extremely close to the physical noise floor dictated by the discreteness of ionic charge.
While averaging over h0 statistically independent sensors could, in principle, continue to reduce noise by h1, this scaling breaks down at high sensor density or over increasing spatial extent. As the sensor coverage increases, the noise cross-correlations induced by Johnson-Nyquist collective modes become dominant, setting a second, geometry- and dielectric-structure-dependent noise floor for the entire cell.
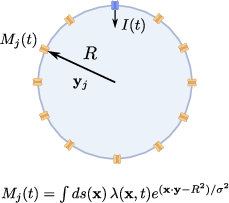
Figure 3: Illustration of a spherical cell with h2 ion channels distributed over its membrane, signaling via surface charge density h3; fluctuations from long-range modes become increasingly important as channel density increases.
Theoretical analysis yields a frequency-dependent crossover ion channel density h4 at which the signal-to-noise achievable through averaging saturates. For biologically relevant timescales:
- For slow signals (h5 ms), h6–h7 channels/µmh8
- For rapid, h9s signaling (e.g., at nodes of Ranvier), this rises to d0–d1 channels/µmd2
These densities are strikingly consistent with direct experimental measurements for neuronal somata, axon initial segments, and nodes of Ranvier.
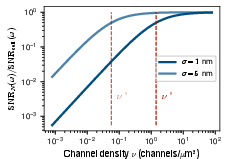
Figure 4: Demonstration of signal-to-noise ratio (SNR) saturation with increasing d3, confirming the theoretical prediction that collective sensing rapidly approaches the “perfect instrument” limit set by Johnson-Nyquist noise.
Local Versus Collective Noise Regimes
A key result is the existence of two distinct and non-interconvertible information limits:
- For d4, averaging can reduce shot noise, producing true statistical gain.
- For d5, all sensors are maximally correlated by collective Johnson-Nyquist modes, so further increasing sensor count yields no new information—the “perfect instrument” limit.
This transition has profound implications: for sufficiently high channel densities, the cell's voltage readout is as accurate as physically possible for the given geometry.
Architectural and Biological Implications
This theory provides biophysical design constraints for both single-channel sensitivity and optimal ion channel deployments for neural computation. Notably, unlike diffusive ligand sensing, the critical number of sensors for Johnson-Nyquist saturation does not scale linearly with cell size, but depends directly on measurement speed and only weakly on spatial dimensions due to collective screening.
Of particular significance: the observed densities of sodium channels in neuronal initiation zones and rapid-conduction regions appear tuned to these physical limits—supporting the hypothesis of evolutionary optimization subject to fundamental noise.
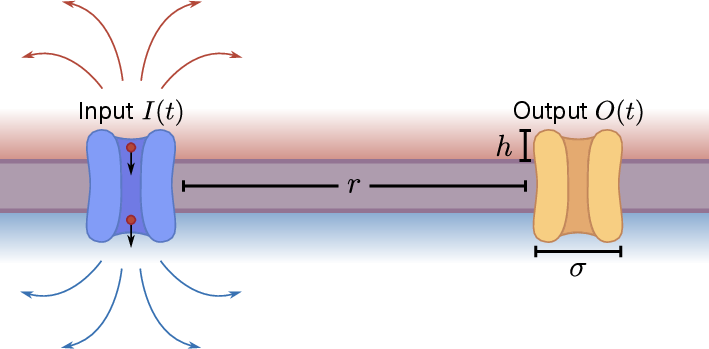
Figure 5: Schematic for signal transmission between two distant ion channels, modeling the propagation of the voltage signal in the presence of both local and global noise constraints.
Prospects for Sensing Architectures
While the analysis focuses on simple charge-integrating sensors, the paper discusses the possibility that alternative architectures—such as sensors based on electric field rather than net charge density—could provide different or superior noise characteristics. However, given the long-range nature of electric fields and the locality of shot noise, it is mathematically nontrivial to overcome the shot noise bound for any channel operating in three-dimensional solution, unless some as-yet-unrealized mechanism is exploited in the specific structure of the voltage sensor.
Future Directions and Theoretical Significance
The identification of thermal fluctuation-imposed limits on neural information processing resonates with classic physical limits of computation and energetics in biological systems. This work implies that even evolved cellular architectures may generically operate in regimes where performance is set not by molecular sophistication, but by the ultimate constraints of equilibrium statistical physics.
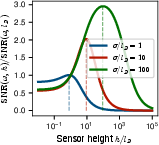
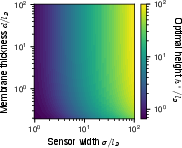
Figure 6: Dependence of SNR on sensor geometry, confirming weak height dependence and underlying the rapid saturation with increasing width/height beyond a few Debye lengths.
Conclusion
This work establishes robust, quantitative bounds on both the speed and precision of voltage sensing by biological ion channels, grounded in first-principles thermodynamics and physical modeling. Numerical results and scaling arguments show that both the architecture of single channel sensors and the density of neural voltage-gated ion channels are tightly constrained by the interplay between discrete shot noise and collective Johnson-Nyquist modes. These findings substantiate the view that neuronal computation is fundamentally bounded by the physics of thermal fluctuations—a result that is both of practical importance for the interpretation of neural physiology and of theoretical significance for the physics of biological information processing.
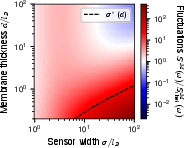
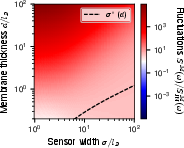
Figure 7: Detailed comparison of measured fluctuations, showing quantitative agreement between theoretical decompositions and numerical analysis, validating the core shot noise versus Johnson-Nyquist scaling picture.
References
For a comprehensive development of the arguments and supporting derivations, see the full article "Thermal fluctuations set fundamental limits on ion channel function" (2604.03538).