- The paper derives explicit analytical solutions for Mi-phase dynamics, demonstrating exponential growth of toxic proteins under defined initial conditions.
- It employs matched asymptotic expansions to connect outer He-phase solutions with inner Mi-phase and logistic To-phase behavior, reinforcing model predictions.
- The study provides practical insights for early intervention and translational applications in neurodegenerative disease modeling.
Analytical Characterisation of Mi- and To-Phases in HeMiTo Dynamics
Introduction
This study presents an in-depth analytical examination of the Mi- (mixed) and To- (toxic) phases within the HeMiTo dynamical framework for prion-like protein propagation. The HeMiTo framework addresses the mechanistic spatiotemporal dynamics of heterodimer-based models that govern misfolded protein aggregation, an essential process implicated in neurodegenerative diseases such as Alzheimer's. Previous results established the outer solution for the initial healthy phase (He-phase) analytically, but provided only numerical evidence for the Mi-phase (exponential onset) and inferred the To-phase (saturation) through stability analysis. This work closes those gaps by deriving explicit inner solutions, performing matched asymptotics, and formalising the reduction to classical growth laws.
The HeMiTo Model Class and Phase Structure
The class of heterodimer ODE models is expressed as:
\begin{align*}
\frac{\mathrm{d}u}{\mathrm{d}\tau} &= c_1 - c_2 u - \varepsilon u v f(v) \
\frac{\mathrm{d}v}{\mathrm{d}\tau} &= \varepsilon v (u f(v) - 1)
\end{align*}
with positive ε≪1, analytic f(v), and biologically feasible initial conditions. Two stationary states (HSS, TSS) are present under generic constraints on f. Perturbation expansions produce outer (He-phase) and inner (Mi-phase) solutions. Previous work had established that uHe(τ) evolves monotonically to the HSS, while v remains small; the Mi-phase initiates when v begins to increase, preceding the To-phase where the system approaches the TSS.
Analytical Solution of Mi-Phase via Inner/Outer Matching
The inner solution for the Mi-phase is derived through systematic expansion in ε and matched to the outer solution of the He-phase. The result is an explicit solution for both healthy (uMi(τ)) and toxic (vMi(τ)) species. The matching determines that the maximum healthy concentration is exactly the HSS value, and the profile of u(τ) is strictly concave—initially increasing and then declining after reaching the maximum at the onset of the Mi-phase.
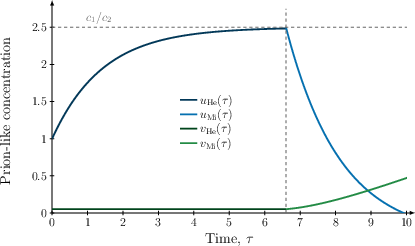
Figure 1: Asymptotic matching of the inner and outer solutions confirms the maximum of the healthy species at the HSS and delineates the transition between He- and Mi-phases.
The Mi-phase solution for the toxic species, f(v)0, admits rigorous approximation by an exponential function for early times:
f(v)1
provided that f(v)2. This inequality gives an exact criterion for the emergence of exponential growth and identifies the mechanistic dependence of the Mi-phase "take-off" on both the kinetic conversion function and initial healthy species abundance.
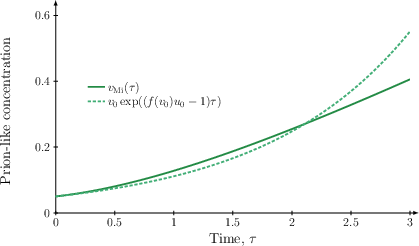
Figure 2: Exponential approximation of the toxic species during the Mi-phase, demonstrating the accuracy of the analytical result for small f(v)3.
This analytical characterisation of Mi-phase dynamics strengthens previous numerical observations, establishing that exponential onset is not artefactual but an inherent property governed by system parameters.
Logistic Saturation in the To-Phase
Proceeding to the To-phase, the analysis invokes a quasi steady-state (QSS) reduction in the neighbourhood of the TSS. The healthy species f(v)4 becomes quasi-stationary, evolving on an f(v)5 time scale, and can be algebraically eliminated:
f(v)6
Substitution into the equation for f(v)7 yields a one-dimensional scalar ODE which, under local expansion, is reducible to a logistic growth law:
f(v)8
with explicit expressions for the growth rate f(v)9 and carrying capacity f0, both expressible in terms of model parameters and the conversion function f1. Critically, the saturation plateau of the toxic species aligns with the TSS, linking the late-stage (To-phase) dynamics to the broader asymptotic behaviour of the system.
Implications and Future Directions
This comprehensive analytical deconstruction of the Mi- and To-phases has multiple theoretical and translational implications:
- Mechanistic Validation: The precise correspondence of exponential and logistic phases with classic biomarker trajectories provides mechanistic underpinning for descriptive clinical models.
- Predictive Utility: The explicit phase-dependent analytical formulae facilitate parameter inference and real-time forecasting when applied to longitudinal biomarker or imaging data, e.g., in disease progression studies.
- Therapeutic Targeting: Exact identification of dynamical regimes can inform strategies targeting early intervention (suppressing exponential onset) versus late-stage plateau management.
- General Applicability: The heterodimer framework encompasses not only prion-like diseases but any system with autocatalytic conversion dynamics, extending its relevance across infectious disease and aggregate biology settings.
Further theoretical developments may focus on extensions to spatially-resolved models (PDEs), stochastic variants, and systems with more complex feedback via f2 or external regulation. Empirically, there is promise for integrating such mechanistic models with state-of-the-art inference toolkits (e.g., PINTS) for patient-specific forecasting.
Conclusion
This work rigorously elucidates the analytical structure of the Mi- and To-phases in the HeMiTo framework for prion-like conversion, deriving explicit phase-wise solutions, transition criteria, and reduction to canonical growth laws. These results unify and mechanistically ground empirical observations of exponential and logistic biomarker trajectories in neurodegeneration and establish a predictive foundation for future translational applications in disease modelling and intervention design (2604.00871).