- The paper demonstrates that interspecific information use relaxes the competitive exclusion principle by altering interaction topologies and enabling stable coexistence.
- It employs an extended consumer-resource model with both deterministic ODEs and stochastic simulations to quantify behavioral and demographic impacts.
- Empirical validations from seabirds, flour beetles, and fruit flies support its findings and underscore its broad ecological applicability.
Introduction
The classical Competitive Exclusion Principle (CEP) posits that multiple consumer species cannot stably coexist on a single limiting resource. Yet, extensive empirical evidence, such as the paradox of the plankton and diverse species assemblages in forests and grasslands, reveals persistent violations of this principle. This paper systematically investigates the role of interspecific information use among predators as a theoretically robust and empirically supported mechanism that fundamentally relaxes CEP constraints, enabling species coexistence even in simple consumer-resource systems.
The analysis extends MacArthur’s consumer–resource framework with explicit behavioral coupling via information use. The system comprises two consumer populations, C1 and C2, and a single resource R. Individual-level dynamics are described by:
{x˙i=aiCi(F)R(F)−(ki+di)xi C˙i=wikixi−DiCi R˙=g({R},{xi},{Ci})(i=1,2)
Parameters ai, ki, and di encode encounter, capture, and escape rates, respectively; wi is the biomass conversion efficiency, Di the mortality rate; resource dynamics g(⋅) follow logistic growth for biotic resources and external influx for abiotic resources.
Interspecific information use is modeled by a Monod-type augmentation of encounter rates:
a1(C2)=a1′[1+K2+C2l2C2]
where l2 controls the maximal efficiency boost and K2 sets the half-saturation for the effect.
Dimensionless analysis ensures scalability and generality. System steady-states and stability are evaluated via a fixed-point analysis with Jacobian eigenvalues, supported by both deterministic ODE and stochastic SSA simulations.
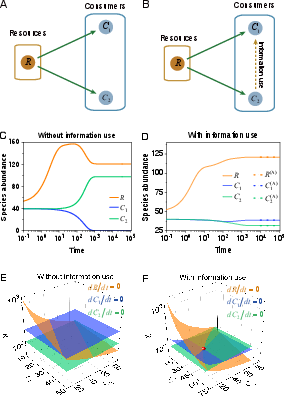
Figure 1: Interspecific information use transforms the phase space geometry, inducing non-parallel zero-growth isoclines and enabling stable fixed points corresponding to coexistence.
Mechanistic Breakdown of the Competitive Exclusion Principle
In the absence of interspecific information use, the zero-growth manifolds for C1 and C2 are parallel planes in (C1,C2,R) space, precluding intersection and stable coexistence. The introduction of interspecific information use via a1(C2) changes the topology: C1’s manifold becomes a function of C2, and the intersection of three non-parallel surfaces emerges, corresponding to a physically realizable coexistence fixed point.
Stable coexistence is not a result of fine-tuning; systematic parameter scans elucidate broad domains in (D1,D2) space where coexistence is robust. Both biotic and abiotic resource cases exhibit stretching of coexistence regions in parameter space, confirmed by eigenvalue analysis and trajectory simulations.
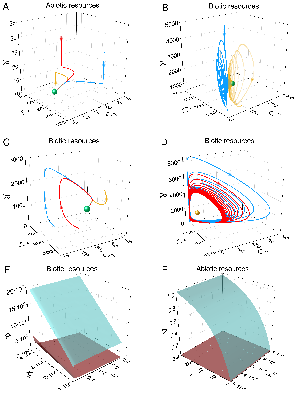
Figure 2: Dynamical coexistence modes include stable fixed points (global attractors) and limit cycles, revealing both steady-state and oscillatory biodiversity maintenance.
Robustness to Stochasticity
To examine ecological validity in fluctuating environments, stochastic simulation algorithms (SSA) are implemented. The coexistence equilibria persist under demographic noise and environmental variability. SSA and ODE results are statistically consistent, evidenced by low rRMSE (<0.18) and high Pearson correlation (>0.997) between simulated and empirical species abundances.
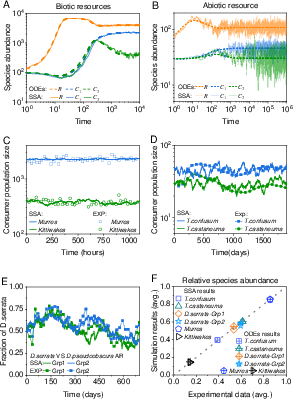
Figure 3: Time courses from stochastic and deterministic simulations align quantitatively with field and laboratory data, supporting ecological realism of the coexistence mechanism.
Application to Empirical Systems and Contradictory Experiments
Three flagship systems are quantitatively accounted for:
- Seabird Assemblages: Black-legged Kittiwakes and Murres cohabiting cliff ecosystems, with behavioral foraging cues facilitating mutual persistence.
- Tribolium Flour Beetles: Long-term coexistence of T. castaneum and T. confusum sharing a single flour resource, potentially enabled by pheromonal information use.
- Drosophila Fruit Flies: Experimental coexistence in mixed-species populations, consistent with behaviorally mediated information transfer.
Model predictions for relative abundances closely match experimental and field survey data (see main text and supplementary figures), confirming the explanatory power of the mechanism.
Theoretical Implications and Generalization
The analytical solutions show that interspecific information use effectively introduces a novel “information resource,” adding degrees of freedom to the coexistence equations. This resource augments the trophic structure: while both consumer species exploit R, one acts as a source of information for the other, analogously to facilitation processes in toxin-mediated or cross-feeding models.
The mechanism is biologically plausible and theoretically generalizable. Prior models based on generalized Lotka–Volterra or omnivory could not permit n consumers to coexist on a single resource due to structural constraints; the present approach circumvents this by explicit modeling of individual-level encounters and behavioral information transfer.
Real-world implications extend to microbial communities (quorum sensing, coordinated defense), multi-trophic systems, and ecosystem management. However, in complex systems with additional metabolic and evolutionary constraints (cross-feeding, substrate partitioning, overflow metabolism), caution is warranted: coexistence may require integration of multiple mechanisms.
Figure 4: Empirical validation across multiple ecological systems establishes mechanistic links between behavioral information use and observed community structure.
Conclusion
This study rigorously demonstrates that interspecific information use among consumers constitutes a robust, general mechanism for breaking the classical constraints of the competitive exclusion principle. By transforming interaction topologies in consumer-resource systems, such information use allows for persistent coexistence, supported by robust analytical, numerical, and empirical analysis. The implications span behavioral ecology, microbial dynamics, and biodiversity theory, offering avenues for integrating behavioral processes into quantitative models of ecosystem stability.
Future directions include extension to multi-species networks, further exploration of higher-order interactions, and incorporation of eco-evolutionary feedbacks in complex ecosystems. Integrative frameworks combining behavioral information, metabolic exchange, and stochasticity are recommended for predictive modeling of biodiversity under global change scenarios.

