Context dependent adaptation in a neural computation
Abstract: Brains adapt to the statistical structure of their input. In the visual system, local light intensities change rapidly, the variance of the intensity changes more slowly, and the dynamic range of contrast itself changes more slowly still. We use a motion-sensitive neuron in the fly visual system to probe this hierarchy of adaptation phenomena, delivering naturalistic stimuli that have been simplified to have a clear separation of time scales. We show that the neural response to visual motion depends on contrast, and this dependence itself varies with context. Using the spike-triggered average velocity trajectory as a response measure, we find that context dependence is confined to a low-dimensional space, with a single dominant dimension. Across a wide range of conditions this adaptation serves to match the integration time to the mean interval between spikes, reducing redundancy.
Paper Prompts
Sign up for free to create and run prompts on this paper using GPT-5.
Top Community Prompts
Explain it Like I'm 14
Plain-language summary of “Context dependent adaptation in a neural computation”
1. What is this paper about?
This paper explores how a single motion-sensing neuron in a fly’s brain automatically adjusts the way it reads visual motion depending on the situation. The key idea is that the neuron doesn’t just react to what’s happening right now (how “contrasty” the image is at this moment), but also to the broader context—what range of contrasts it has been seeing recently. The authors show that this “context-dependent adaptation” helps the neuron use its spikes efficiently so that each spike carries fresh, non-redundant information.
2. What did the researchers want to find out?
They asked simple versions of these questions:
- How does a motion-sensing neuron’s response change when the contrast (how much images vary between light and dark) goes up or down?
- Does the response depend only on the current contrast, or also on the bigger picture—the range of contrasts the fly has been seeing over the last minutes (the “context”)?
- Can these changes be described simply, or are they complicated?
- Do these adjustments help the neuron avoid repeating itself (sending multiple spikes that say the same thing)?
3. How did they study it?
They recorded spikes from a well-studied fly neuron called H1, which detects horizontal motion. To test adaptation cleanly, they built visual movies with a clear separation of time scales:
- Making the scene: They showed the fly a textured, natural-looking pattern and moved it randomly left-right so the “speed” (velocity) changed very fast.
- Controlling contrast: They slowly changed how strongly light and dark patches differed (the “contrast,” like turning the sharpness up or down). This slow contrast was the “instantaneous” condition the neuron saw.
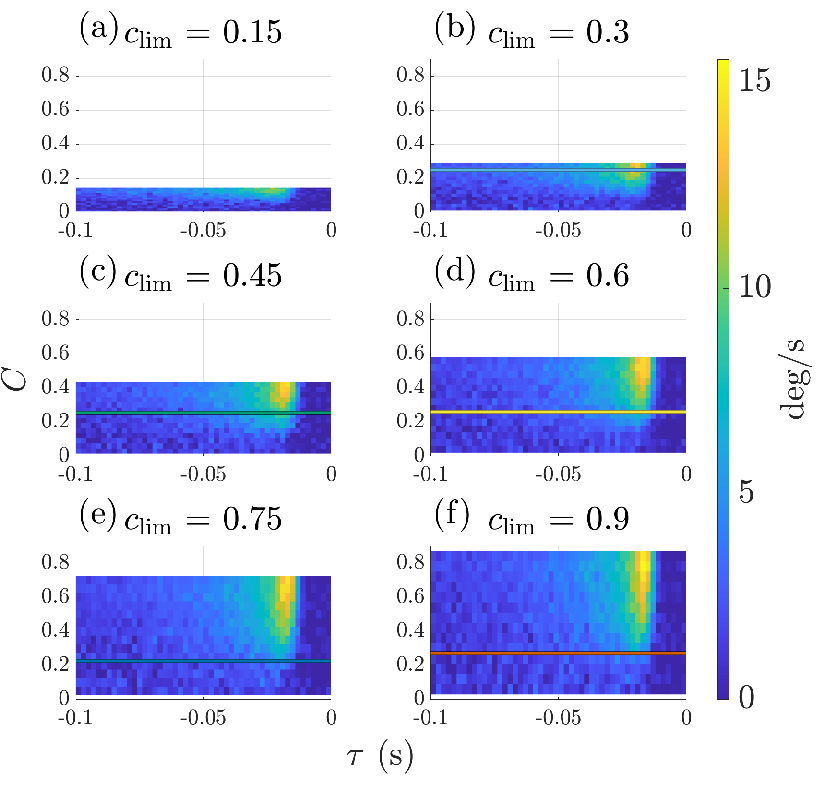
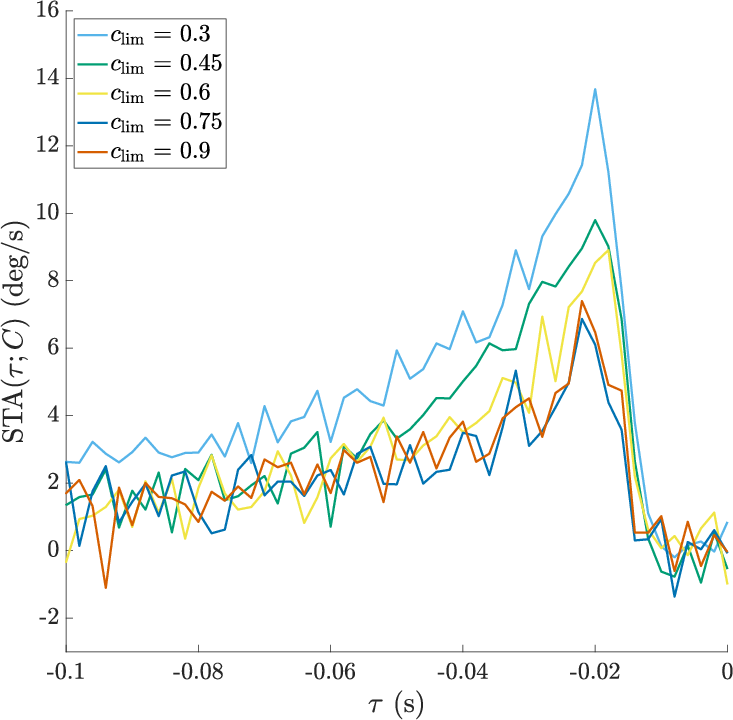
- Setting context: They grouped experiments by the overall range of contrasts allowed (from narrow to wide). That range is the “context”—like telling the system, “In this session, contrasts will vary only a little,” or “In this session, they may vary a lot.”
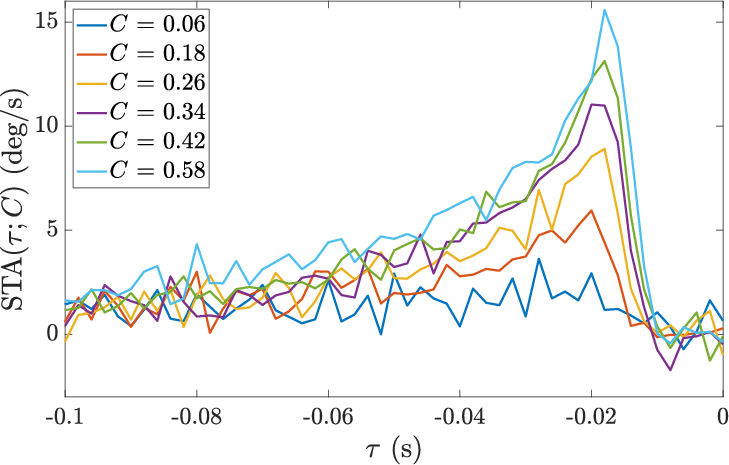
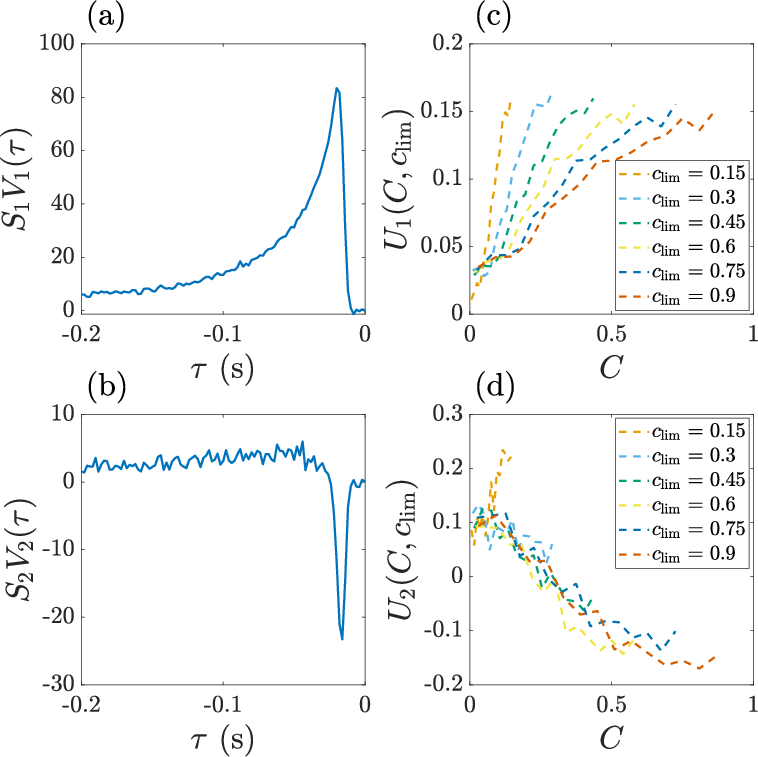
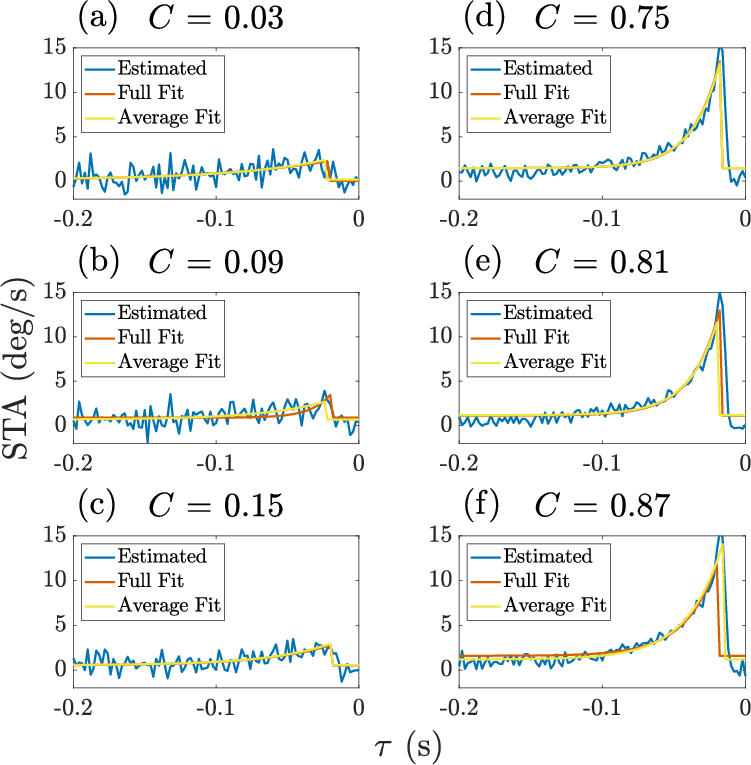
- Measuring the neuron’s motion response: They used the spike-triggered average (STA). Think of this as the “average mini-movie of speed” right before each spike. If you line up many spikes and average the motion that came before them, you see the typical motion pattern that makes the neuron fire. They did this separately for different contrast levels and different contexts.
- Finding the main patterns: They looked for the simplest way to describe how all these STAs change with contrast and context. In everyday terms, this is like finding the top two or three “themes” that explain most of the changes, instead of needing to track every tiny detail.
Key idea in plain terms:
- Spike-triggered average (STA): Like rewinding a few frames before every “beep” the neuron makes and averaging those frames to see the typical speed pattern that triggers the beep.
- Context: Not just the current picture’s contrast, but the recent “rules of the game” about how big contrasts can get.
- Main patterns analysis: Like discovering most songs on an album share the same beat and melody, and you only need those few ingredients to explain the whole album.
4. What did they find, and why does it matter?
Main results:
- The neuron’s motion response depends on both current contrast and context.
- Higher contrast made the neuron more sensitive to motion and shortened how long it “listens” (its integration time). That makes sense: when the picture is clear, you don’t need to listen for long to be confident.
- But the same contrast value produced different responses in different contexts. For example, “25% contrast” triggered a stronger or weaker response depending on whether the overall session allowed only small or very large contrasts. So the neuron doesn’t just look at “now”; it also adjusts to what it has been seeing lately. That’s context-dependent adaptation.
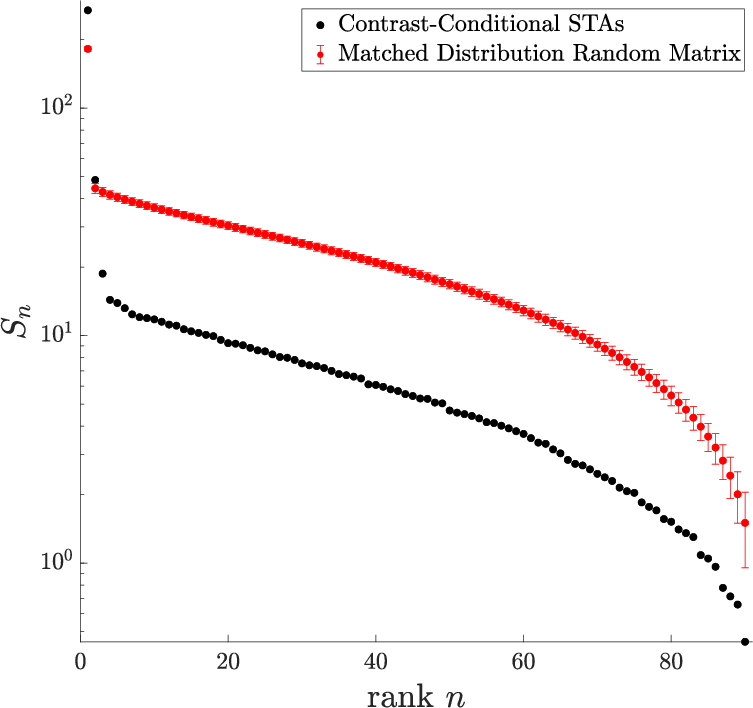
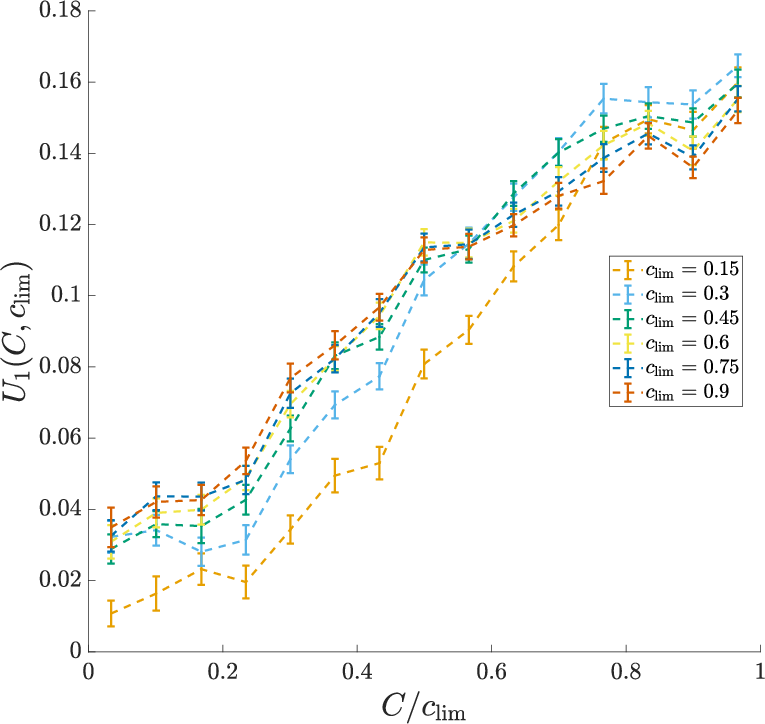
- Simple structure: Despite many possible ways the response could change, almost everything was explained by just two main patterns (a low-dimensional, rank-2 description).
- The dominant pattern mostly depended on “contrast divided by the context’s range” (current contrast scaled by how big contrasts have been recently). This is like normalizing the current signal to the size of the whole playing field.
- A second, smaller pattern depended on absolute contrast (the raw level), almost independent of context. This may help resolve situations when the signal is small and noise matters.
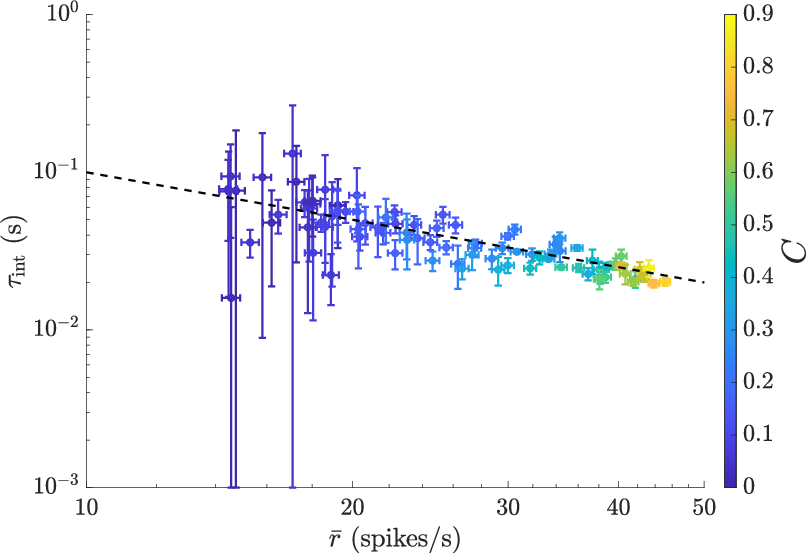
- Timing is tuned to reduce redundancy: The neuron’s “listening window” (integration time) was about the same as the average time between spikes. In other words, τ_integration ≈ 1 / spike rate.
- Why is this interesting? If the neuron listened much longer than the gap between spikes, different spikes would contain overlapping information about the same bit of motion—wasting spikes by repeating the message. Matching the two time scales helps each spike carry fresh information.
- They observed this across many contrasts and contexts—suggesting the neuron actively keeps itself out of a “rate coding” regime where you need many spikes to say one thing. Instead, it aims for “one spike per meaningful moment.”
Why this matters:
- It shows a neuron can adapt not only to “what’s happening now” but also to “what usually happens around here.” This is a hallmark of efficient coding—saving resources while keeping important information.
- The discovery that adaptation fits into just two main patterns means the brain may use simple rules to manage complex inputs.
5. What could this mean going forward?
- For neuroscience: The brain seems to constantly match its computations to the statistics of the world—normalizing signals to their recent range and tuning timing so each spike matters. This supports the idea that nervous systems aim for efficient, low-redundancy codes.
- For other senses and species: Similar context-sensitive strategies may be common in seeing, hearing, and touch across animals.
- For technology: Cameras and sensors could borrow this trick—automatically scaling their sensitivity to the recent range of inputs and adjusting how long they “listen” so each data point adds new information.
- For understanding behavior: Efficient spikes likely help fast, reliable behaviors (like flight control in flies), where quick, non-repetitive messages are crucial.
In short, this study shows that a fly’s motion neuron constantly adjusts how strongly and how long it listens to the world, based on both the current clarity of the scene and the broader context of recent visual experience. It keeps its messages short, timely, and informative—so every spike counts.
Collections
Sign up for free to add this paper to one or more collections.

