Noise robustness and metabolic load determine the principles of central dogma regulation
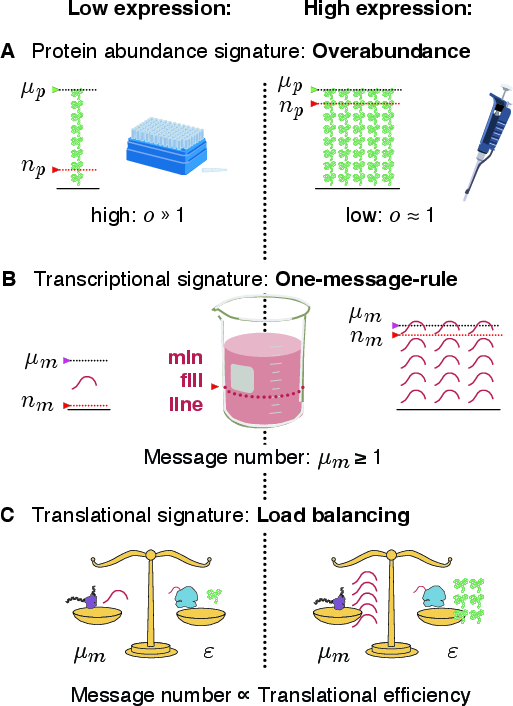
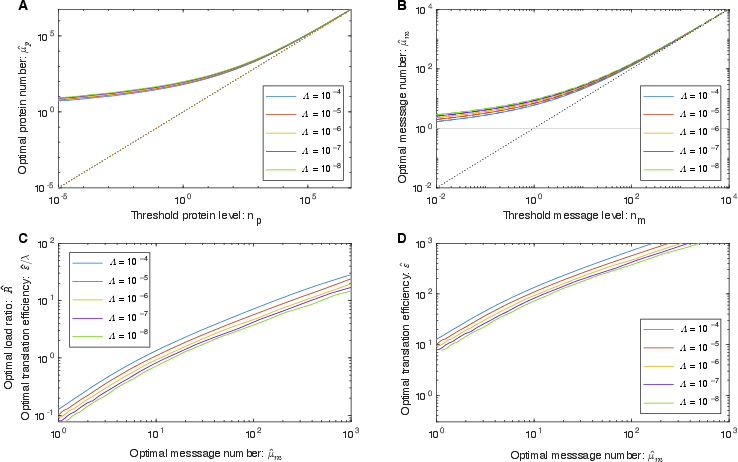
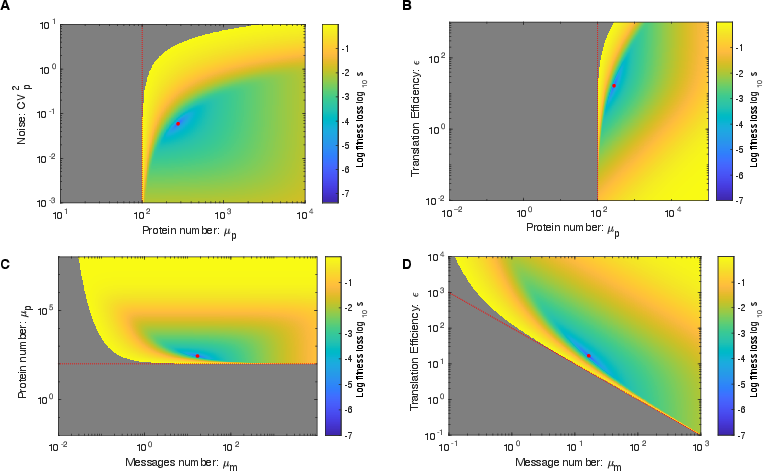
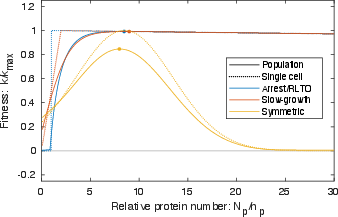
Abstract: The processes of gene expression are inherently stochastic, even for essential genes required for growth. How does the cell maximize fitness in light of noise? To answer this question, we build a mathematical model to explore the trade-off between metabolic load and growth robustness. The model predicts novel principles of central dogma regulation: Optimal protein expression levels for many genes are in vast overabundance. Essential genes are transcribed above a lower limit of one message per cell cycle. Gene expression is achieved by load balancing between transcription and translation. We present evidence that each of these novel regulatory principles is observed. These results reveal that robustness and metabolic load determine the global regulatory principles that govern gene expression processes, and these principles have broad implications for cellular function.
- Nature 436, 588-92 (2005).
- Cell 166, 1282-1294.e18 (2016).
- Cell 165, 1493-1506 (2016).
- Proc Natl Acad Sci U S A 117, 18010-18017 (2020).
- Cell Syst 12, 924-944.e2 (2021).
- Science 309, 2010-3 (2005).
- Nature 441, 840-6 (2006).
- Science 329, 533-8 (2010).
- Nat Commun 10, 68 (2019).
- F. Crick, Central dogma of molecular biology. Nature 227, 561-3 (1970).
- Phys Rev Lett 84, 5447-50 (2000).
- Phys Rev Lett 97, 168302 (2006).
- Science 330, 1099-102 (2010).
- Phys. Rev. Lett. 107, 218101 (2011).
- mBio 12, e0256121 (2021).
- Cell Syst 12, 56-67.e6 (2021).
- Mol Syst Biol 16, e9828 (2020).
- Science 350, 1096-101 (2015).
- Nature 425, 737-41 (2003).
- Science 378, eabk2066 (2022).
- Nature 473, 337-42 (2011).
- PLoS Biol. 19, e3001416 (2021).
- Nat Commun 8, 1225 (2017).
- PLoS Genet 10, e1004556 (2014).
- Biochim Biophys Acta 9, 648-60 (1952).
- Cell 184, 4579-4592.e24 (2021).
- Phys Rev E 105, 014420 (2022).
- FASEB J 3, 2360-70 (1989).
- J Biol Chem 217, 947-57 (1955).
- Cell Syst 5, 283-294.e5 (2017).
- D. Gillespie, A rigorous derivation of the chemical master equation. Physica A 188, 404–425 (1992).
- D. T. Gillespie, Exact stochastic simulation of coupled chemical reactions. The Journal of Physical Chemistry 81, 2340–2361 (1977).
- Proc Natl Acad Sci U S A 99, 9697-702 (2002).
- Mol Syst Biol 11, 808 (2015).
- (2023).
- FEMS Yeast Res 16 (2016).
- Nat Commun 14, 1762 (2023).
- Cell Rep 14, 22-31 (2016).
- G3 (Bethesda) 10, 3229-3242 (2020).
- Philos Trans A Math Phys Eng Sci 374 (2016).
- R. Milo, What is the total number of protein molecules per cell volume? A call to rethink some published values. Bioessays 35, 1050-5 (2013).
- Mol Gen Genet 170, 137-44 (1979).
- PLoS One 5, e15442 (2010).
- Mol Cell Biol 19, 7357-68 (1999).
- Genome Res 13, 1863-72 (2003).
- Nucleic Acids Res 38, D750-3 (2010).
- Methods Mol Biol 416, 183-94 (2008).
- Sci Rep 9, 11005 (2019).
- Cell 10, 453-62 (1977).
- T. von der Haar, A quantitative estimation of the global translational activity in logarithmically growing yeast cells. BMC Syst Biol 2, 87 (2008).
- Nat Struct Mol Biol 15, 1263-71 (2008).
- BMC Genomics 9, 574 (2008).
- Nature 455, 1251-4 (2008).
- Science 347, 1260419 (2015).
- Nat Genet 23, 387-8 (1999).
- J Bacteriol 185, 5673-84 (2003).
- mBio 9 (2018).
- BMC Microbiol 4, 4 (2004).
- Nat Genet 38, 636-43 (2006).
- Science 297, 1183-6 (2002).
- Proc Natl Acad Sci U S A 99, 12795-800 (2002).
- C. R. Rao, Information and the accuracy attainable in the estimation of statistical parameters. Bulletin of Calcutta Mathematical Society 37, 81-91. (1945).
- Cell Syst 4, 495-504.e5 (2017).
- Scandinavian Journal of Statistics 41, 1051–1063 (2014).
Paper Prompts
Sign up for free to create and run prompts on this paper using GPT-5.
Top Community Prompts
Collections
Sign up for free to add this paper to one or more collections.