- The paper presents a novel finding showing that confined epithelial monolayers exhibit power-law, scale-free velocity correlations.
- It uses micropatterned PDMS substrates and high-resolution KLT tracking to precisely quantify directional migration and velocity fields.
- Results indicate that substrate stiffness and cytoskeletal perturbations significantly influence velocity coordination, advancing models of tissue mechanics.
Scale-Free Velocity Correlations in Confined Epithelial Cell Monolayers
Introduction
The mechanical and collective migration properties of epithelial tissues are essential for processes such as embryogenesis, wound repair, and cancer metastasis. While significant progress has been made in elucidating the roles of cell-intrinsic activity, cell–cell interactions, and substrate mechanics, the emergence and scaling of velocity correlations within confined cell monolayers remain incompletely described. The paper "Geometric Confinement Reveals Scale-Free Velocity Correlations in Epithelial Cell Monolayer" (2511.21655) presents a rigorous experimental investigation into the interplay of geometry, topology, and substrate rigidity in dictating spatiotemporal velocity correlations in MDCK II epithelial monolayers, with an emphasis on the breakdown of classical length-scale-dependent behavior in favor of scale-free, power-law correlations.
Experimental Techniques and Geometric Paradigms
The authors deploy micropatterned PDMS substrates, enabling precise control of adhesive regions and mechanical boundary conditions. Two principal geometries are probed:
- Strip with Free Front: A long open-ended strip with a central circular non-adhesive obstacle, supporting directed migration following removal of a mobile boundary.
- Closed Racetrack: A millimeter-scale annular (racetrack-shaped) adhesive pattern, optionally containing obstacles but devoid of any free front, implemented on both hard (MPa) and soft (kPa) PDMS to probe rigidity effects.
Mitomycin C treatment suppresses proliferation, circumventing jamming effects and enabling monolayer migration and pattern maturation over extended timescales (tens of hours), with high temporal (∼5 min) and spatial (∼10 μm) resolution imaging. Velocity fields are extracted using the Kanade-Lucas-Tomasi (KLT) feature tracking algorithm.
Influence of Geometric Boundaries and Substrate Mechanics
Strips: The free front establishes a robust velocity gradient, driving bulk flows oriented toward the leading edge and modulating both velocity magnitudes and density distributions. Local perturbations due to the non-adhesive obstacle reveal anisotropic flow deviation, with pronounced acceleration and deceleration zones demarcated laterally and longitudinally with respect to the obstacle.

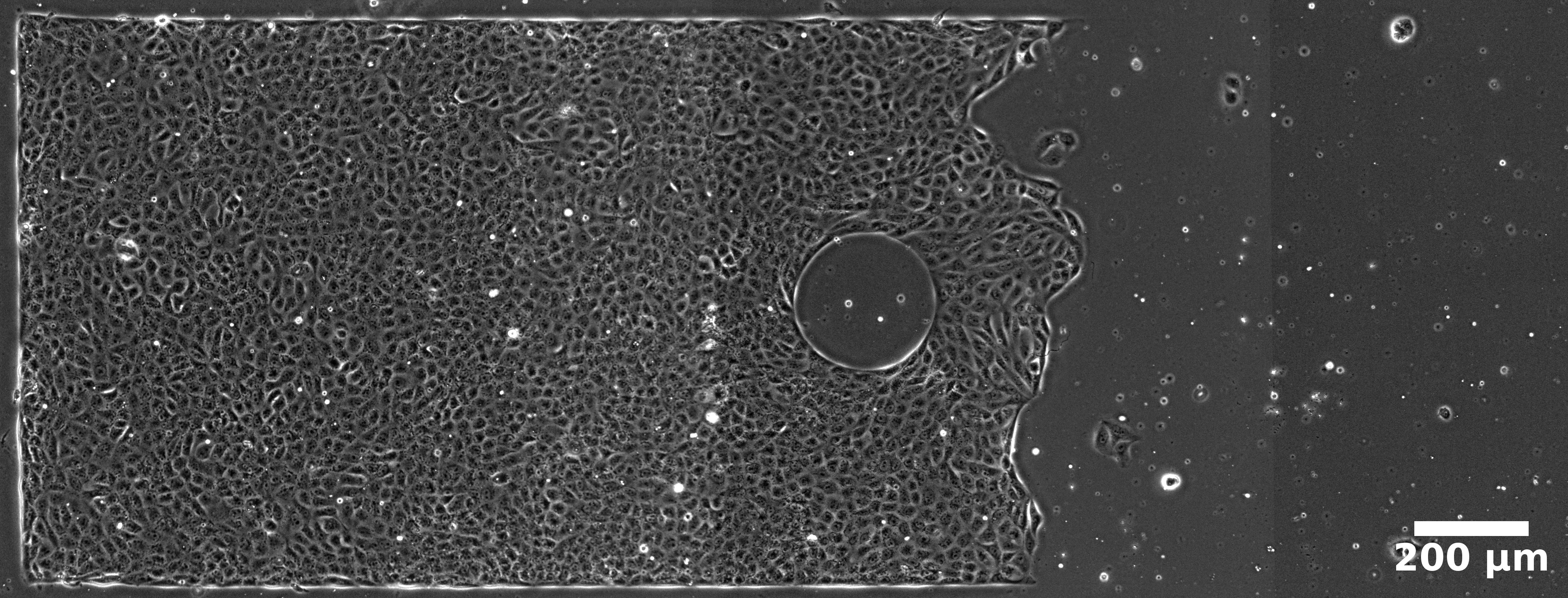

Figure 1: Velocity field in a strip with a central obstacle, showing dominant directed flow and anisotropic perturbation domains.
Vimentin staining demonstrates that biochemical polarization cues extend significantly further from free fronts (characteristic decay length 65±4 μm) compared with confining edges (3.2±0.7 μm), implicating front-induced gradients in cytoskeletal remodeling far into the monolayer.
Racetracks: In the absence of a free front, collective migration is not dominated by a global drift. Instead, the velocity field displays competing domains of spontaneous circulation, typically with zero mean flow over long timescales. The introduction of obstacles drives only localized circulation, and the baseline velocity variance and coordination depend on substrate stiffness.


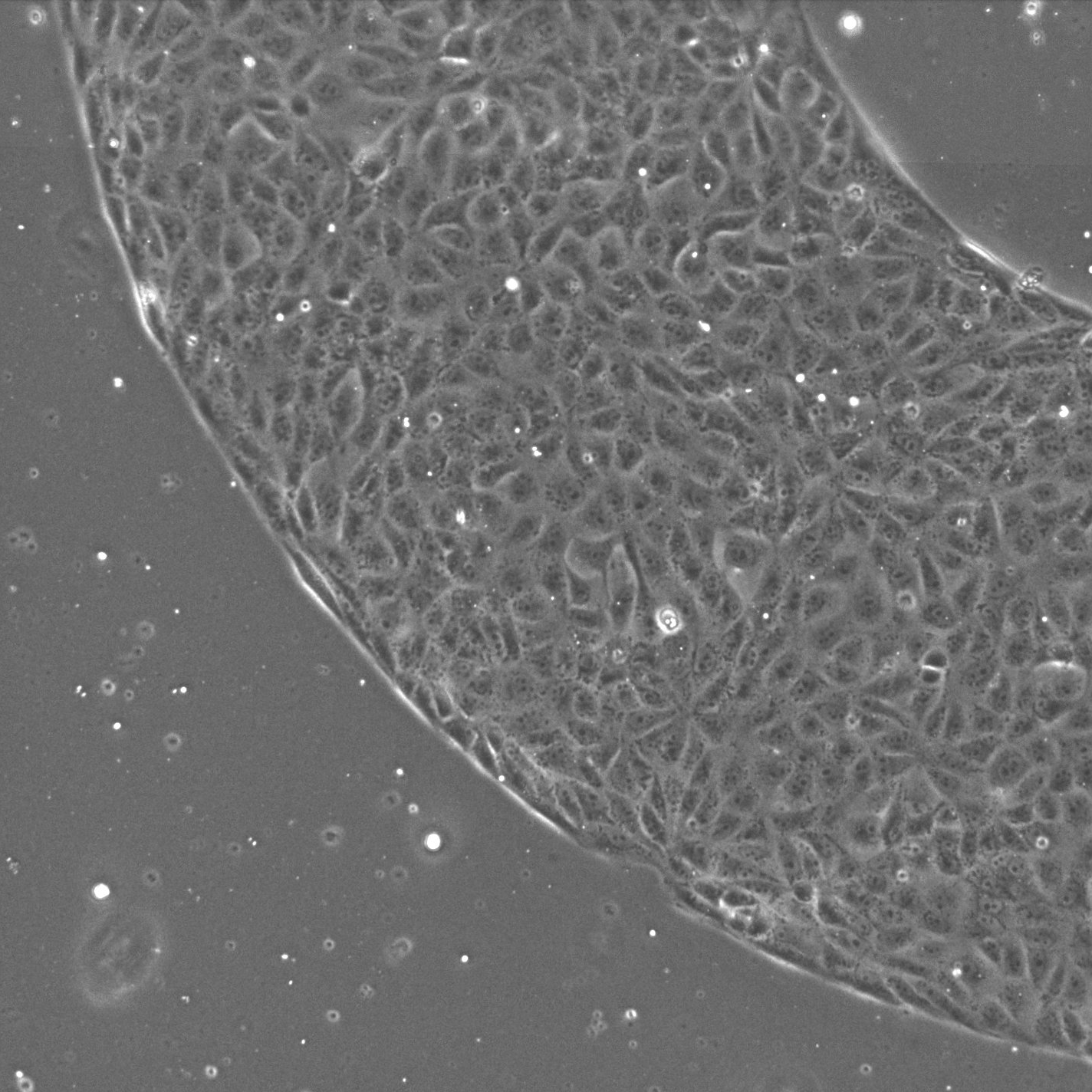

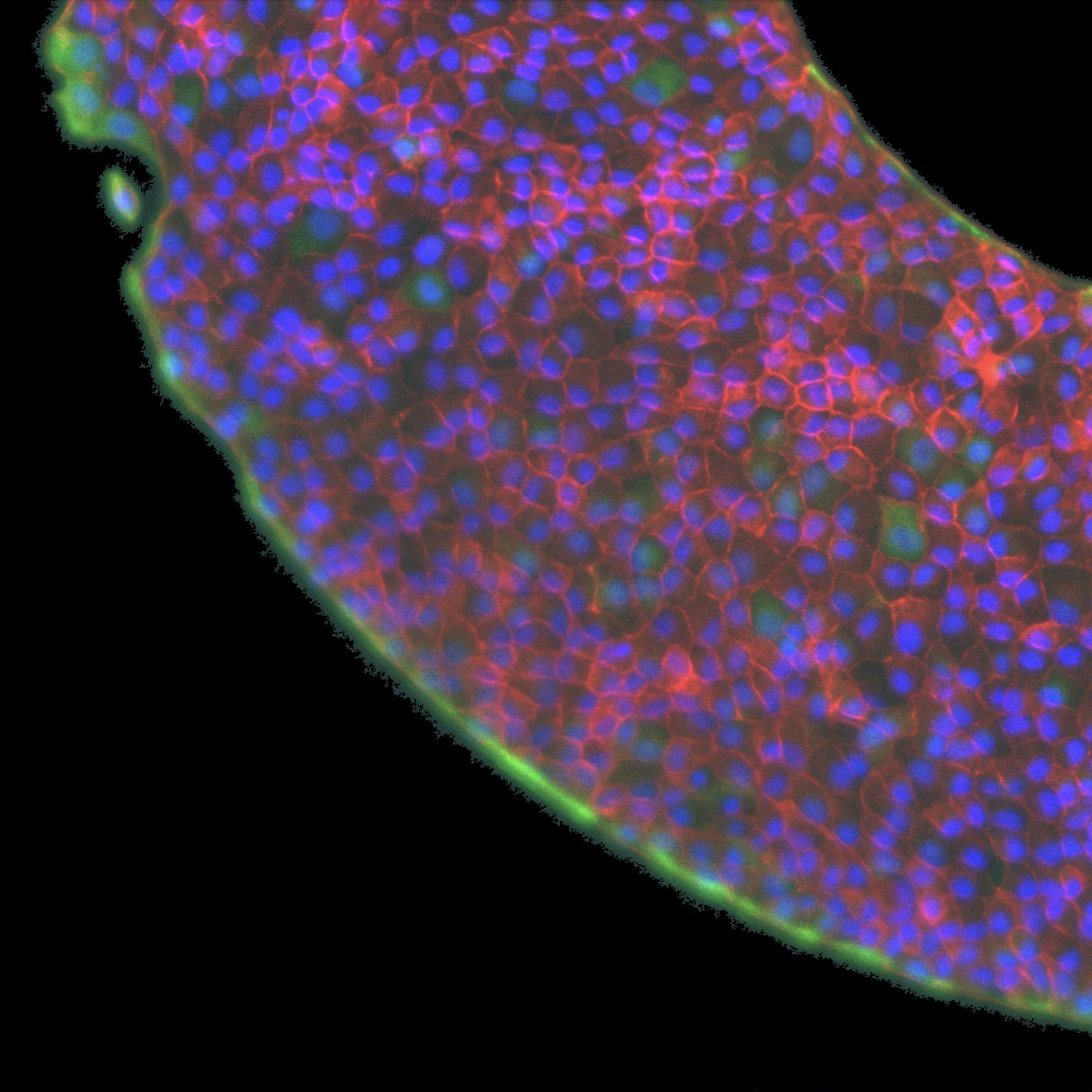
Figure 3: Racetrack setup, illustrating the closed geometry with adhesive tracks, confining boundaries, and embedded obstacles.
Traction force microscopy on the soft racetrack reveals that traction peaks at confining edges align with vimentin-rich domains, supporting the functional linkage between mechanical stress transmission and intermediate filament biochemistry.
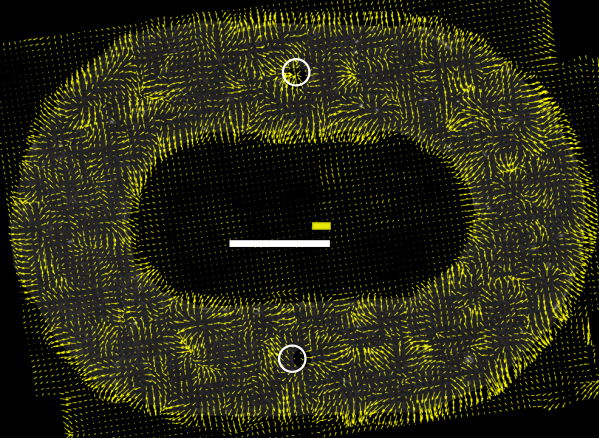

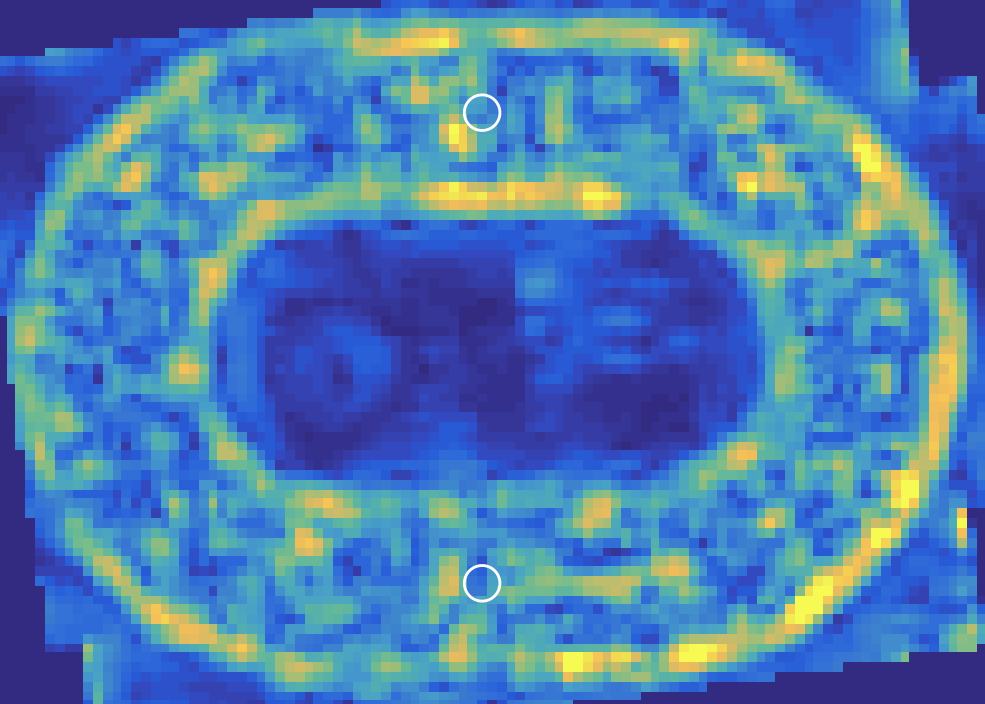


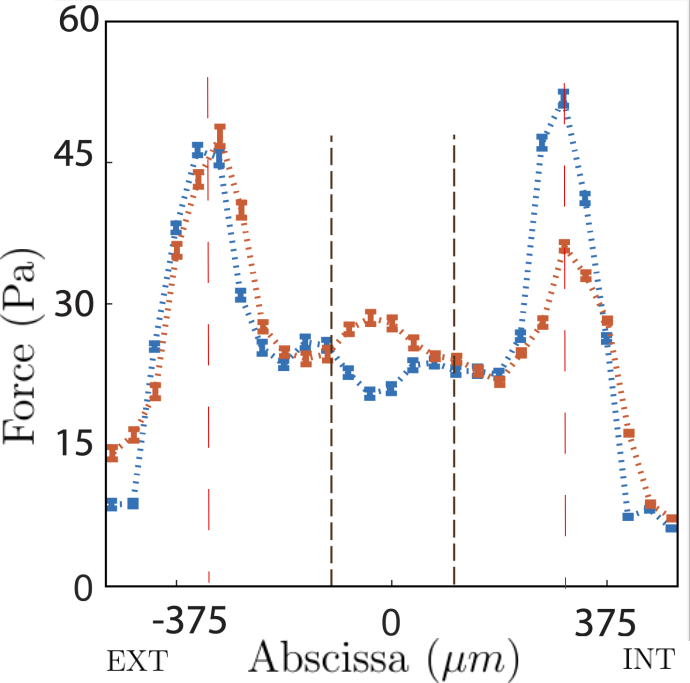
Figure 5: Traction force map for a soft racetrack, highlighting boundary-localized tractions and their spatial correspondence with vimentin staining.
A key quantitative result is that average migration velocity and coordination parameters are substantially higher on soft substrates, with velocity moduli and their standard deviation increasing by up to 50% and 30% respectively (Student's p<0.01 for both measures).
Emergence of Scale-Free Velocity Correlations
Spatial velocity autocorrelation functions were computed in both geometries but analytically focused on racetracks due to the absence of sustained macroscopic gradients. Contrary to the hypothesis of a unique, intrinsic correlation length governing collective cell motion, the authors report:
- Power-law velocity correlations across scales up to the system size, Cs(r)∼rα, with α evolving during monolayer maturation.
- The exponent α is sensitive to cytoskeletal perturbations: actin and microtubule disruption (via CK666, Nocodazole) and vimentin network destabilization (Simvastatin) induce sizable changes, lowering α toward −1.
- No evidence is found for a plateau or characteristic domain size; instead, the patterning wavelength and domain coherence scale non-monotonically with system width and time.


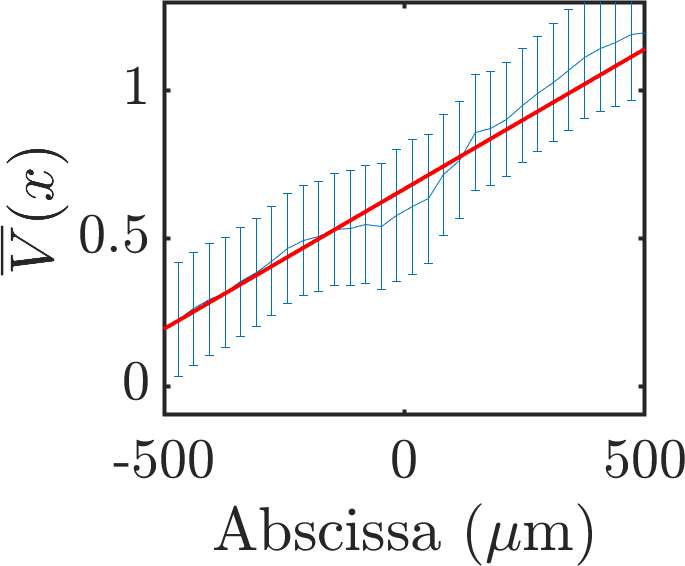
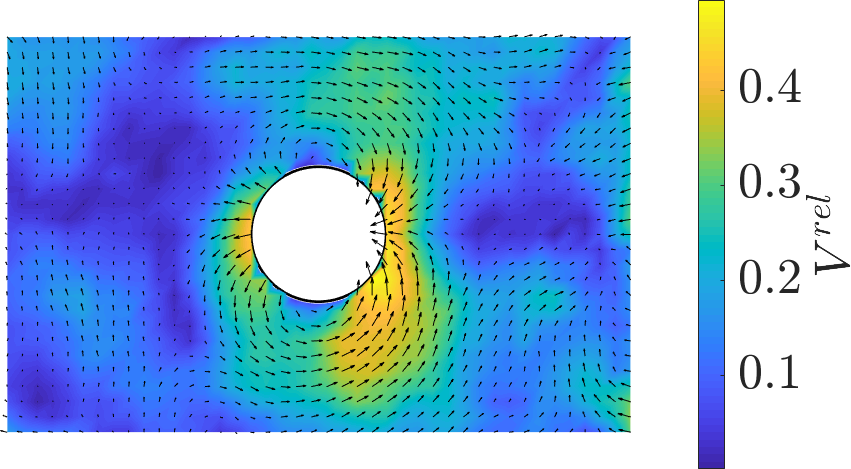
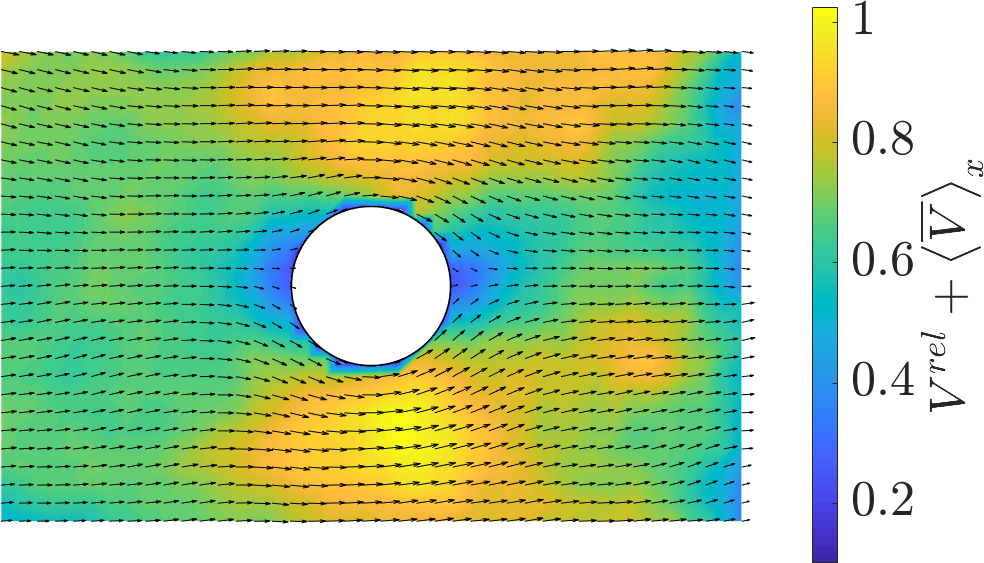
Figure 2: Velocity domain structure along the racetrack midline, quantified by autocorrelation minima, showing system-size scaling and long-range coherence.
The temporal correlation function for velocity fluctuations decays exponentially with a characteristic time constant of ∼100 min, invariant across geometric constraints.
Figure 4: Traction forces on the soft racetrack highlight mechanical boundary effects and simultaneous vimentin enrichment.
Energetic and Vortical Signatures
Analysis of kinetic energy and enstrophy distributions reveals Boltzmann-like statistics and non-monotonic temporal evolution. Both quantities increase during initial domain growth, then energy plateaus while enstrophy decays; these observations are indicative of the emergence and decay of mesoscale vortices ("swirls") inherent to active nematic systems without jamming constraints.

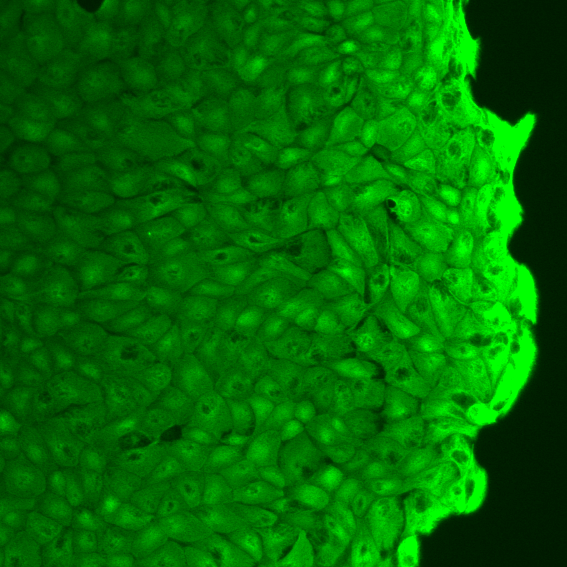
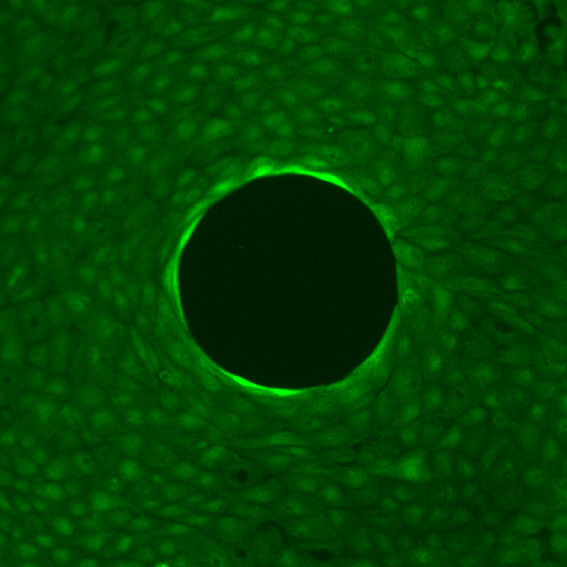
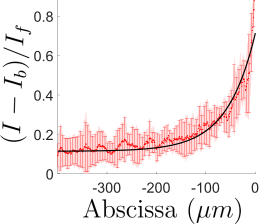
Figure 6: Kinetic energy and enstrophy statistics show Boltzmann distributions and non-monotonic evolution consistent with transient formation and dissipation of large-scale vortical flow.
Implications and Future Directions
The paper demonstrates that, under confinement and in the absence of free fronts, epithelial monolayers exhibit velocity correlations lacking a well-defined length scale and instead governed by system-spanning power laws. This contradicts prior assumptions of a single, intrinsic coordination scale and suggests the collective migration of confluent tissues is best conceptualized in the context of critical-like, scale-free active solids.
The sensitivity of correlation exponents to cytoskeletal perturbation positions these metrics as potent probes of tissue mechanical state and as a bridge between molecular-scale interventions and emergent tissue rheology.
From a theoretical perspective, these results motivate extensions of active matter and glassy dynamics models to explicitly incorporate geometric boundary conditions, long-range stress propagation, and temporal maturation. Practically, the linkage between confinement, substrate mechanics, and mechanical signaling (e.g., vimentin gradients) has implications for the design of biomimetic scaffolds and control of migration in tissue engineering contexts.
Conclusion
By systematically varying geometry, topology, and rigidity, this study provides a comprehensive characterization of collective velocity statistics in MDCK epithelial tissues. The discovery of robust, scale-free velocity correlations imposes a revision of paradigms for active tissue mechanics under confinement. This work highlights the necessity of large-scale, long-duration experiments to capture critical phenomena and offers new quantifiable observables (power-law exponents) for interpreting and controlling collective cell migration. The findings also set the stage for integrative multi-scale modeling that links cytoskeletal modulation, mechanical stress fields, and emergent active matter behavior in tissues.