On Conformal Isometry of Grid Cells: Learning Distance-Preserving Position Embedding
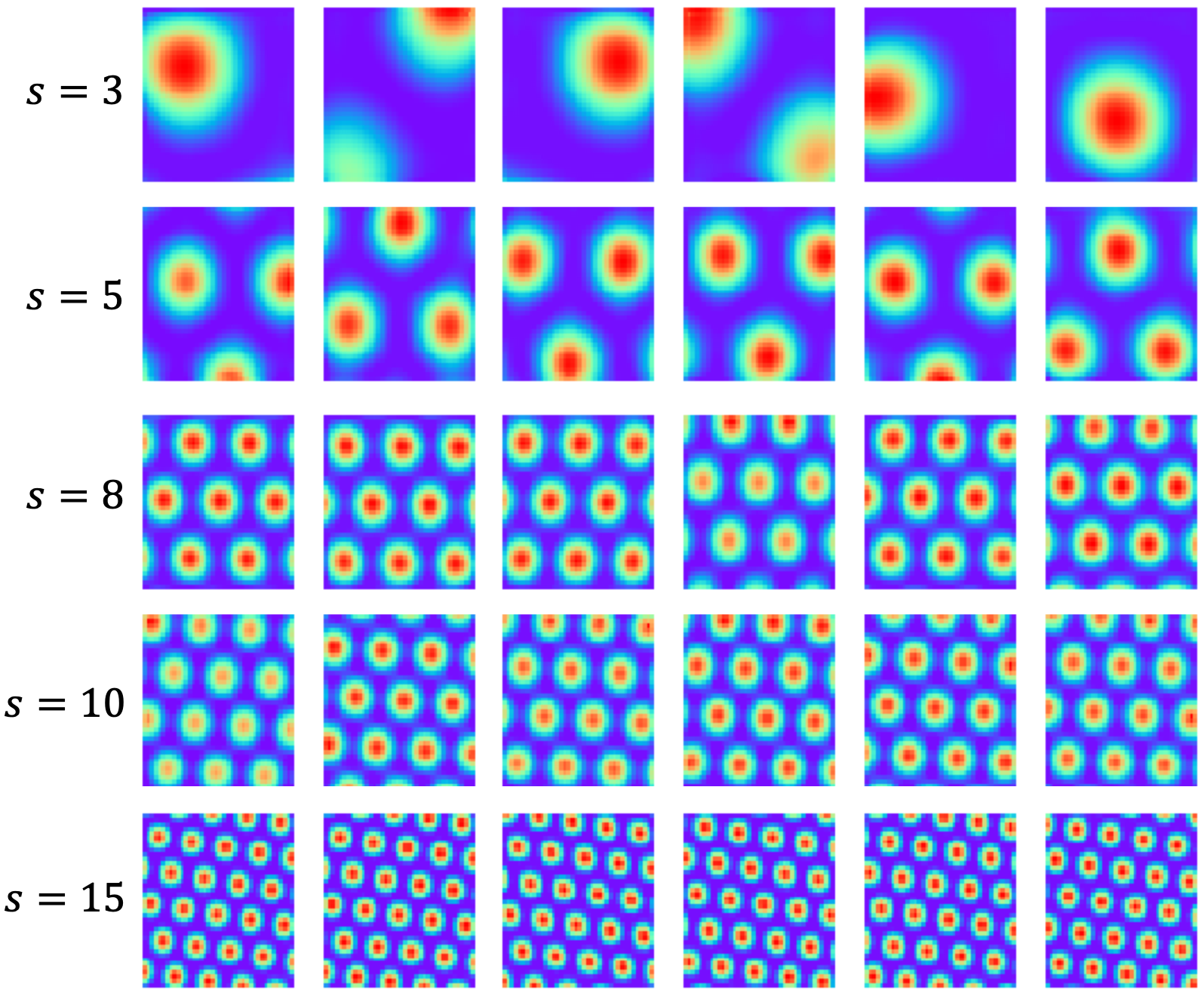
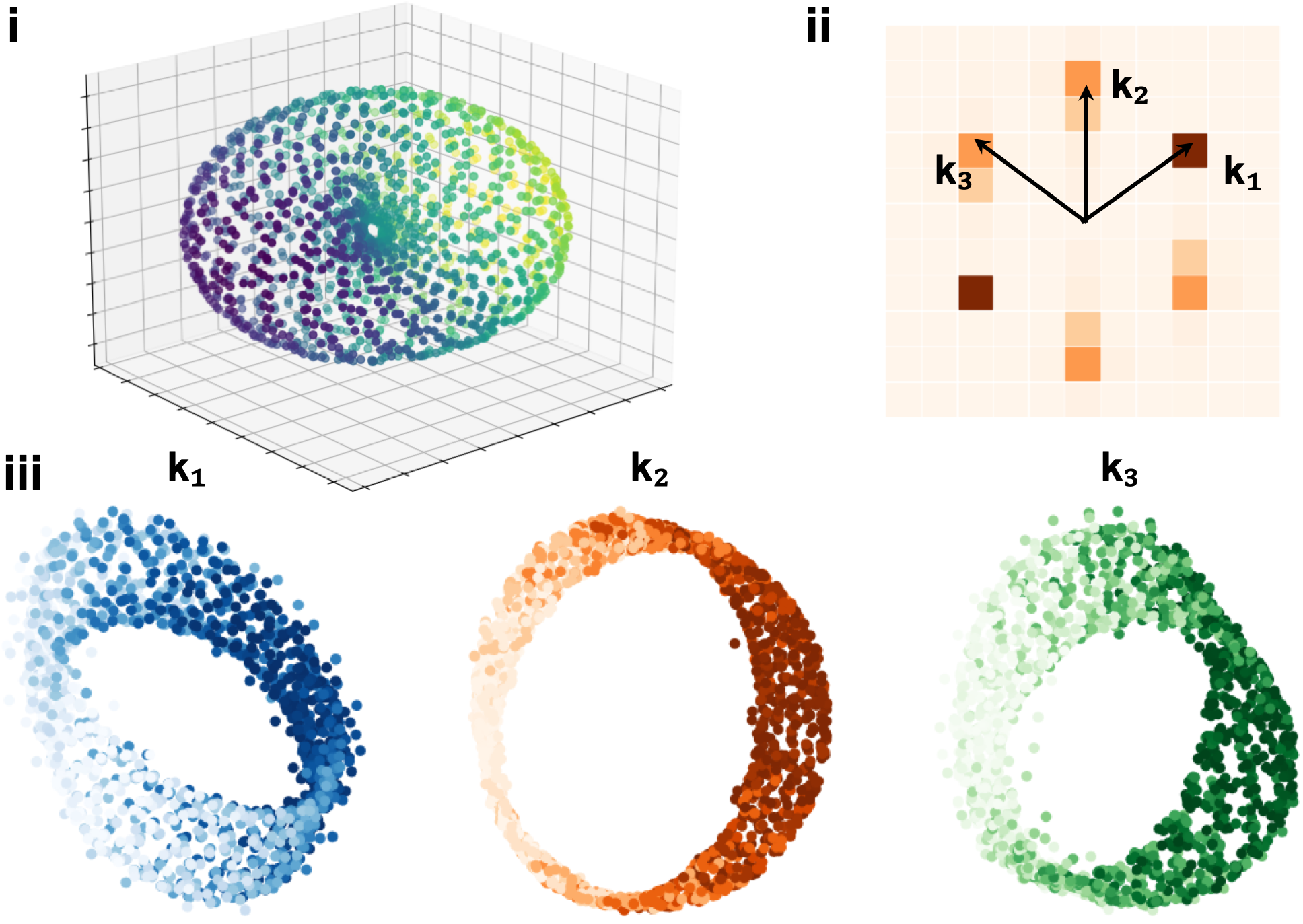
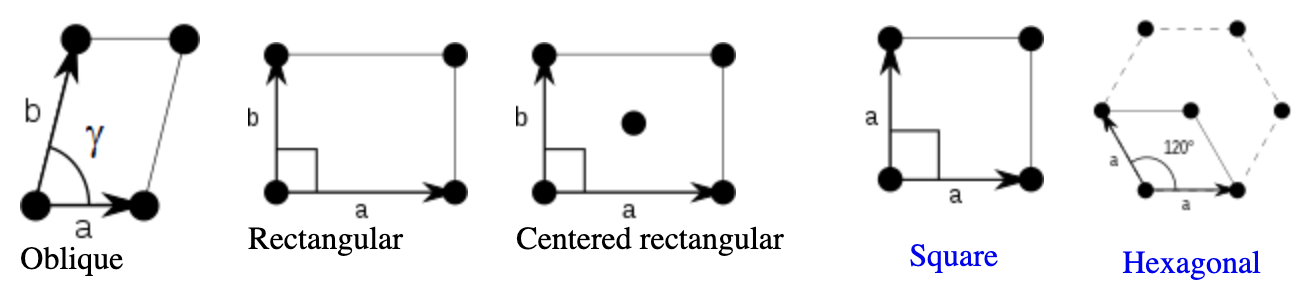
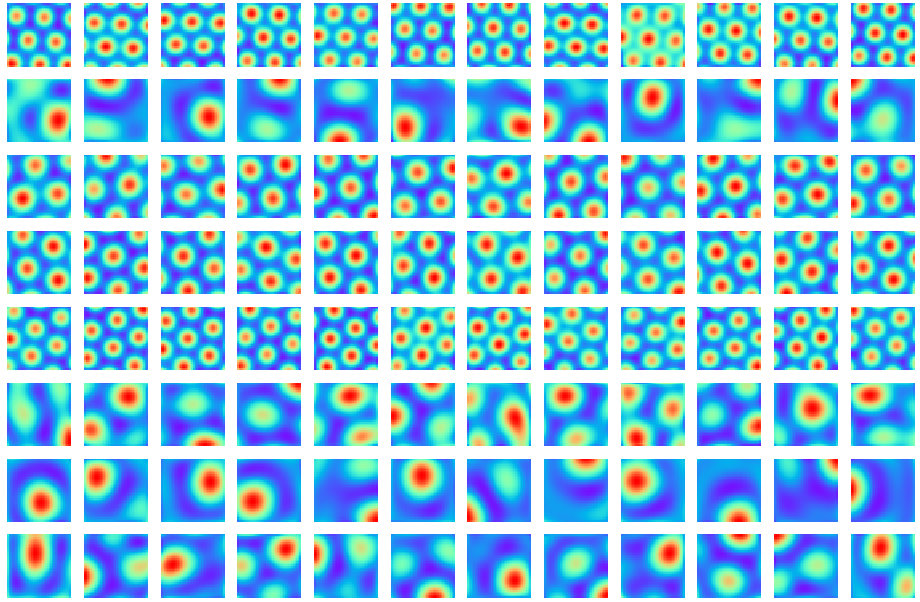
Abstract: This paper investigates the conformal isometry hypothesis as a potential explanation for the hexagonal periodic patterns in grid cell response maps. We posit that grid cell activities form a high-dimensional vector in neural space, encoding the agent's position in 2D physical space. As the agent moves, this vector rotates within a 2D manifold in the neural space, driven by a recurrent neural network. The conformal hypothesis proposes that this neural manifold is a conformal isometric embedding of 2D physical space, where local physical distance is preserved by the embedding up to a scaling factor (or unit of metric). Such distance-preserving position embedding is indispensable for path planning in navigation, especially planning local straight path segments. We conduct numerical experiments to show that this hypothesis leads to the hexagonal grid firing patterns by learning maximally distance-preserving position embedding, agnostic to the choice of the recurrent neural network. Furthermore, we present a theoretical explanation of why hexagon periodic patterns emerge by minimizing our loss function by showing that hexagon flat torus is maximally distance preserving.
- A theory of joint attractor dynamics in the hippocampus and the entorhinal cortex accounts for artificial remapping and grid cell field-to-field variability. eLife, 9:e56894, 2020.
- Daniel J Amit. Modeling brain function: The world of attractor neural networks. Cambridge university press, 1992.
- Solid state physics, 1976.
- Vector-based navigation using grid-like representations in artificial agents. Nature, 557(7705):429, 2018.
- Experience-dependent rescaling of entorhinal grids. Nature neuroscience, 10(6):682–684, 2007.
- Navigating cognition: Spatial codes for human thinking. Science, 362(6415):eaat6766, 2018.
- Scale-invariant memory representations emerge from moire interference between grid fields that produce theta oscillations: a computational model. Journal of Neuroscience, 27(12):3211–3229, 2007.
- The entorhinal cognitive map is attracted to goals. Science, 363(6434):1443–1447, 2019.
- Accurate path integration in continuous attractor network models of grid cells. PLoS computational biology, 5(2):e1000291, 2009.
- Organizing conceptual knowledge in humans with a gridlike code. Science, 352(6292):1464–1468, 2016.
- Recurrent inhibitory circuitry as a mechanism for grid formation. Nature neuroscience, 16(3):318–324, 2013.
- Emergence of grid-like representations by training recurrent neural networks to perform spatial localization. arXiv preprint arXiv:1803.07770, 2018.
- Emergence of functional and structural properties of the head direction system by optimization of recurrent neural networks. International Conferences on Learning Representations (ICLR), 2020.
- The input–output transformation of the hippocampal granule cells: from grid cells to place fields. Journal of Neuroscience, 29(23):7504–7512, 2009.
- Evidence for grid cells in a human memory network. Nature, 463(7281):657, 2010.
- Extracting grid cell characteristics from place cell inputs using non-negative principal component analysis. Elife, 5:e10094, 2016.
- Actionable neural representations: Grid cells from minimal constraints. arXiv preprint arXiv:2209.15563, 2022.
- William Gerard Dwyer and CW Wilkerson. The elementary geometric structure of compact lie groups. Bulletin of the London Mathematical Society, 30(4):337–364, 1998.
- What grid cells convey about rat location. Journal of Neuroscience, 28(27):6858–6871, 2008.
- A spin glass model of path integration in rat medial entorhinal cortex. Journal of Neuroscience, 26(16):4266–4276, 2006.
- Spatial representation in the entorhinal cortex. Science, 305(5688):1258–1264, 2004.
- Grid cells in mice. Hippocampus, 18(12):1230–1238, 2008.
- Learning grid cells as vector representation of self-position coupled with matrix representation of self-motion. In International Conference on Learning Representations, 2019.
- On path integration of grid cells: group representation and isotropic scaling. In Neural Information Processing Systems, 2021.
- Toroidal topology of population activity in grid cells. bioRxiv, 2021.
- Impaired path integration in mice with disrupted grid cell firing. Nature neuroscience, 21(1):81–91, 2018.
- Are grid cells used for navigation? on local metrics, subjective spaces, and black holes. Neuron, 111(12):1858–1875, 2023.
- Microstructure of a spatial map in the entorhinal cortex. Nature, 436(7052):801, 2005.
- Grid-like processing of imagined navigation. Current Biology, 26(6):842–847, 2016.
- Direct recordings of grid-like neuronal activity in human spatial navigation. Nature neuroscience, 16(9):1188, 2013.
- A map of visual space in the primate entorhinal cortex. Nature, 491(7426):761, 2012.
- Adam: A method for stochastic optimization. arXiv preprint arXiv:1412.6980, 2014.
- Development of the spatial representation system in the rat. Science, 328(5985):1576–1580, 2010.
- Path integration and the neural basis of the’cognitive map’. Nature Reviews Neuroscience, 7(8):663, 2006.
- Where am i? where am i going? Scientific American, 314(1):26–33, 2016.
- Explaining heterogeneity in medial entorhinal cortex with task-driven neural networks. Advances in Neural Information Processing Systems, 34:12167–12179, 2021.
- The hippocampus as a spatial map: preliminary evidence from unit activity in the freely-moving rat. Brain research, 1971.
- Précis of o’keefe & nadel’s the hippocampus as a cognitive map. Behavioral and Brain Sciences, 2(4):487–494, 1979.
- Feedback inhibition enables theta-nested gamma oscillations and grid firing fields. Neuron, 77(1):141–154, 2013.
- Impaired speed encoding is associated with reduced grid cell periodicity in a mouse model of tauopathy. bioRxiv, page 595652, 2019.
- Functional properties of stellate cells in medial entorhinal cortex layer ii. Elife, 7:e36664, 2018.
- Conjunctive representation of position, direction, and velocity in entorhinal cortex. Science, 312(5774):758–762, 2006.
- Spectral methods for dimensionality reduction. Semi-supervised learning, 3, 2006.
- No free lunch from deep learning in neuroscience: A case study through models of the entorhinal-hippocampal circuit. Advances in Neural Information Processing Systems, 35:16052–16067, 2022.
- Self-supervised learning of representations for space generates multi-modular grid cells. arXiv preprint arXiv:2311.02316, 2023.
- Coherently remapping toroidal cells but not grid cells are responsible for path integration in virtual agents. bioRxiv, pages 2022–08, 2022.
- Hexagons all the way down: Grid cells as a conformal isometric map of space. bioRxiv, pages 2024–02, 2024.
- A unified theory for the origin of grid cells through the lens of pattern formation. Advances in neural information processing systems, 32, 2019.
- A unified theory for the computational and mechanistic origins of grid cells. Neuron, 111(1):121–137, 2023.
- Grid cells generate an analog error-correcting code for singularly precise neural computation. Nature neuroscience, 14(10):1330, 2011.
- The hippocampus as a predictive map. Nature neuroscience, 20(11):1643, 2017.
- Connecting multiple spatial scales to decode the population activity of grid cells. Science Advances, 1(11):e1500816, 2015.
- The entorhinal grid map is discretized. Nature, 492(7427):72, 2012.
- Edward C Tolman. Cognitive maps in rats and men. Psychological review, 55(4):189, 1948.
- Attention is all you need. Advances in neural information processing systems, 30, 2017.
- A principle of economy predicts the functional architecture of grid cells. Elife, 4:e08362, 2015.
- Relating transformers to models and neural representations of the hippocampal formation. arXiv preprint arXiv:2112.04035, 2021.
- Conformal isometry of lie group representation in recurrent network of grid cells. arXiv preprint arXiv:2210.02684, 2022.
- Grid cells without theta oscillations in the entorhinal cortex of bats. Nature, 479(7371):103, 2011.
- Optogenetic dissection of entorhinal-hippocampal functional connectivity. Science, 340(6128), 2013.
Paper Prompts
Sign up for free to create and run prompts on this paper using GPT-5.
Top Community Prompts
Collections
Sign up for free to add this paper to one or more collections.

